
UNIDAD 3
1
ESTRUCTURA DEL APARATO
RESPIRATORIO
Normalmente con el término respiración
se define el intercambio de gases entre el medio ambiente externo y el medio
interno. Sin embargo, bajo esta definición tan simple se incluye no solamente
el movimiento de aire entre el interior y exterior de los pulmones, sino
también el paso de los mismos del interior pulmonar a la sangre; el transporte
mediante la vía sanguínea hasta las células y su posterior difusión a través de
las membranas celulares. Todos estos pasos permiten a las células el consumo de
O2 y la liberación de CO2. Desde un punto de vista más limitado, como es
el celular, la respiración (o respiración celular) se refiere al metabolismo
oxidativo (oxidación de nutrientes) para la generación de energía metabólica; y
en este proceso es dónde se consume el oxígeno y se forma anhídrido carbónico.
Para poder realizar todas las
funciones descritas se requiere la participación de otros aparatos además del
respiratorio. Así el aparato cardiovascular o la sangre son piezas tan
importantes e imprescindibles como el propio aparato respiratorio.
Funciones no
respiratorias del aparato respiratorio
Además del intercambio gaseoso, el
aparato respiratorio desarrollas otras funciones. Así:
·
El
lecho capilar pulmonar actúa como un filtro para la sangre, ya que pequeños
coágulos, restos celulares o burbujas de aire son eliminados en este aparato.
·
Las
vías aéreas ejercen una gran acción de defensa del organismo, impidiendo la
entrada de agentes patógenos en el cuerpo.
·
Participa
en mecanismos homeostáticos como el control de la temperatura, control de
líquidos corporales, control ácido-básico, etc.
·
El
lecho capilar pulmonar es un importante reservorio de sangre.
·
Tiene
importantes acciones metabólicas.
Estructura del
aparato respiratorio
El aparato respiratorio se divide en
dos partes desde el punto de vista funcional
a) Sistema de conducción o vías
aéreas.
b) Sistema de intercambio o
superficie alveolar.
Vías
respiratorias o sistema respiratorio conductor
Vías aéreas altas: fosas nasales
y faringe.
Vías aéreas bajas: laringe,
tráquea y bronquios.
1.1
FOSAS NASALES
El aire debe recorrer un largo camino
por tu cuerpo para completar el proceso de la respiración. El primer tramo que
recorre está formado por las fosas nasales, esas dos cavidades alargadas que
observas ubicadas en medio de tu cara, con dos pares de aberturas, unas
anteriores y otras posteriores. Las primeras están situadas en la nariz, y se
mantienen en contacto con el exterior. Las segundas, llamadas coanas, comunican
con el interior.
Las paredes de las fosas nasales están
recubiertas por una mucosa, denominada pituitaria, que presenta tres
protuberancias, conocidas como cornetes. Cuando el aire pasa por este sector,
es entibiado por la gran superficie mucosa del tabique nasal y de los cornetes,
siguiendo su calentamiento durante el paso por las vías respiratorias hasta
llegar a los bronquios, con una temperatura adecuada que no produzca ningún
tipo de efecto nocivo.
1.2
FARINGE
Este tubo musculoso se encuentra ubicado
en el cuello y revestido de membrana mucosa; conecta la nariz y la boca con la
tráquea y el esófago. Por la faringe pasan tanto el aire como los alimentos,
por lo que forma parte tanto del aparato digestivo como del aparato
respiratorio.
A continuación de las fosas nasales
nos encontramos con la faringe, que tiene la característica de ser un segmento
común al sistema respiratorio y al sistema digestivo.
Se extiende desde la base del cráneo
hasta la sexta vértebra cervical. De 13 centímetros de largo, se divide en tres
partes: porción nasal o rinofaringe; porción bucal u orofaringe; y porción
laríngea o laringofaringe.
La rinofaringe o nasofaringe, se
encuentra detrás de la fosas nasales y por sobre el nivel del paladar
membranoso. Excepto este último, sus paredes no tienen movimiento, lo que
significa que su cavidad jamás podrá obstruirse. En esta región se acumula un
tejido linfático, muy desarrollado en los niños, llamado amígdalas
nasofaríngeas, que cuando sufren de hipertrofismo -crecen mucho- reciben el
nombre de adenoides.
Por su parte, la orofaringe se
encuentra limitada arriba por el paladar y abajo por el borde superior de la
epiglotis, que es un cartílago que cubre la abertura de la laringe cuando
comes, evitando así que el alimento entre en la tráquea. Contiene las amígdalas
palatinas, dos masas de tejido linfoide ubicadas en las paredes laterales de la
porción bucal de la faringe.
Las amígdalas son de tamaño variable,
y muchas veces son víctimas de inflamaciones, lo que conocemos como amigdalitis.
La laringofaringe es la parte inferior
de esta cadena, y se extiende desde el borde superior de la epiglotis hasta el
borde inferior del cartílago cricoide.
Como puedes ver, la faringe es solo un
lugar de paso, y tiene una estructura acorde a su función, ya que está
revestida por una capa mucosa que se encarga de atrapar las partículas de polvo
que llegan a este lugar, y que son expulsadas a través de la tos o, en el peor
de los casos, tragadas.
La etapa faríngea de la deglución es
un acto absolutamente reflejo e involuntario. Durante uno o dos segundos, la
respiración se inhibe o frena para dar paso a este proceso; sin embargo, al ser
una acción involuntaria, no alcanzas a darte cuenta de que por un momento has
dejado de respirar.
1.1
LA LARINGE
Su pared está formada por una serie de
cartílagos (tiroides, cricoides, aritenoides, cuneiformes, corniculados o de
Santorini y epiglotis), que se articulan entre sí y poseen distintas formas.
Todos se mueven gracias a la acción de músculos, cuyo interior está recubierto
por una mucosa.
La principal función de la laringe es
generar la voz. Además, deja pasar el aire hacia la tráquea y durante la
deglución le cierra el camino a cuerpos extraños.
Este órgano tiene tres partes:
- Supraglotis o vestíbulo: es el sector
donde está la epiglotis.
- Glotis o parte media: es el lugar
donde se encuentran los pliegues o cuerdas vocales, que son dos bandas de
tejido muscular compuestas por los llamados pliegues verdaderos y falsos. Los
pliegues verdaderos, que encierran la glotis (órgano de la fonación), son los
que producen el sonido de la voz cuando el aire que sale de los pulmones pasa a
las cuerdas vocales haciéndolas vibrar. Los pliegues falsos solo cierran la
entrada a la laringe.
- Infraglotis o parte inferior: está
ubicada entre los pliegues vocales y la tráquea.
1.2
LA TRÁQUEA

Es la vía aérea principal de los
pulmones y se divide en dos grandes ramas llamadas bronquios, que canalizan el
aire a uno de los dos pulmones.
Bajando por la laringe nos
encontramos con la tráquea, un tubo cartilaginoso y membranoso
que mide entre 10 y 11 centímetros con un diámetro igual al de tu dedo índice.
Sus paredes son bastante resistentes, gracias a los 20 anillos cartilaginosos
que posee. Aproximadamente la mitad de la tráquea se encuentra en
el cuello y la otra mitad en el tórax. A la altura
delesternón -ese hueso en forma de espada que tienes en la mitad de la
caja torácica- se divide en dos bronquios, uno derecho y otro izquierdo,
que se dirigen hacia los pulmones.
La tráquea está internamente
recubierta por una capa de mucosa, que es una continuación de la que se
halla en la laringe, y su superficie está revestida de una película
de moco, en el cual se adhieren las partículas de polvo que han logrado
atravesar las vías respiratorias superiores. Este moco no sólo retiene el
polvo, sino que además actúa como bactericida.
ÁRBOL
BRONQUIAL
A partir de la tráquea nacen los
bronquios. Estos se abren en dos ramas que penetran en cada uno de tus
pulmones, junto con vasos sanguíneos y nervios; son estas ramificaciones las
que reciben el nombre deárbol bronquial. Al entrar en los pulmones se producen
varias bifurcaciones a medida que los bronquios se hacen más estrechos. Estas
ramitas más delgadas del árbol, de sólo un milímetro de anchura, son lo
que conocemos como bronquiolos.
Los bronquios cumplen también una función motora. Cuando inspiras, el árbol bronquial se ensancha y alarga, lo que facilita la circulación del aire hacia los alvéolos. Además, también se preocupan de colaborar con la acción de los cilios que se encuentran en la mucosa para evitar que entren partículas extrañas a tus pulmones, todo esto mediante un movimiento de las paredes bronquiales.
Los bronquios cumplen también una función motora. Cuando inspiras, el árbol bronquial se ensancha y alarga, lo que facilita la circulación del aire hacia los alvéolos. Además, también se preocupan de colaborar con la acción de los cilios que se encuentran en la mucosa para evitar que entren partículas extrañas a tus pulmones, todo esto mediante un movimiento de las paredes bronquiales.
1.1
LOS PULMONES
Estos órganos son los
protagonistas del proceso de respiración. Se encuentran en la caja torácica, a
ambos lados del corazón, separados por el mediastino, nombre que recibe el
espacio entre cada uno de ellos.
El aire, luego de pasar por las fosas
nasales, circula por la faringe y llega a la tráquea, que se divide en dos
bronquios, cada uno de los cuales penetra en un pulmón. Los pulmones son los
órganos de la respiración donde se produce la hematosis, proceso durante el
cual los glóbulos rojos absorben oxígeno y se liberan del anhídrido carbónico.
Protegidos por las costillas, se encuentran en la caja torácica, a ambos lados
del corazón, separados por el mediastino, nombre que recibe el espacio entre
cada uno de ellos.
Parecidos a un par de esponjas, forman
uno de los órganos más grandes de tu cuerpo. Su función esencial, compartida
con el sistema circulatorio, es la distribución de oxígeno y el intercambio de
gases. Tienen la capacidad de aumentar de tamaño cada vez que inspiras y de
volver a su tamaño normal cuando el aire es expulsado.
¿Sabías que el pulmón derecho es más grande que el izquierdo? Esto, porque está dividido en tres lóbulos -superior, medio e inferior- y el izquierdo solamente en dos - superior e inferior. Cada uno de los lóbulos se divide en un gran número de lobulillos, en cada uno de los cuales irá a parar un bronquiolo, que a su vez se divide en unas cavidades llamadas vesículas pulmonares; estas forman otras cavidades llamadas alvéolos.
El pulmón está recubierto por una membrana serosa que presenta dos hojas, una que se adhiere a los pulmones, llamada pleura visceral, y otra que tapiza el interior de la cavidad torácica, denominada pleura parietal. Estas dos capas se encuentran en contacto, deslizándose una sobre otra cuando tus pulmones se dilatan o contraen. Entre ellas se encuentra la cavidad pleural, que se encarga de almacenar una pequeña cantidad de líquido, cumpliendo una función lubricadora. Pero la misión principal de la membrana pleural es evitar que tus pulmones rocen directamente con la pared interna de la cavidad torácica, manteniendo una presión negativa que impide el colapso de los pulmones.
¿Sabías que el pulmón derecho es más grande que el izquierdo? Esto, porque está dividido en tres lóbulos -superior, medio e inferior- y el izquierdo solamente en dos - superior e inferior. Cada uno de los lóbulos se divide en un gran número de lobulillos, en cada uno de los cuales irá a parar un bronquiolo, que a su vez se divide en unas cavidades llamadas vesículas pulmonares; estas forman otras cavidades llamadas alvéolos.
El pulmón está recubierto por una membrana serosa que presenta dos hojas, una que se adhiere a los pulmones, llamada pleura visceral, y otra que tapiza el interior de la cavidad torácica, denominada pleura parietal. Estas dos capas se encuentran en contacto, deslizándose una sobre otra cuando tus pulmones se dilatan o contraen. Entre ellas se encuentra la cavidad pleural, que se encarga de almacenar una pequeña cantidad de líquido, cumpliendo una función lubricadora. Pero la misión principal de la membrana pleural es evitar que tus pulmones rocen directamente con la pared interna de la cavidad torácica, manteniendo una presión negativa que impide el colapso de los pulmones.
En estas cavidades alargadas que están
en tu nariz es por donde se intercambian oxígeno y dióxido de carbono con el
medio aéreo
Pleura
Es una membrana serosa que tapiza los
pulmones doblada sobre sí misma. Dispone de dos hojas, la externa o parietal,
adherida a la cara interna de la pared costal; y la interna o visceral, que se
encuentra adherida firmemente a los pulmones. Entre ellas prácticamente no hay
separación, tan sólo un poco de líquido que las mantiene aún más adheridas
entre sí.
El espacio pleural (también denominado
intra o interpleural) separa ambas pleuras unas 5-10 μ y está relleno de unos
20 ml de líquido pleural, obtenidos por ultrafiltración del plasma, que se
están renovando continuamente. Este espacio intrapleural es virtual, pero
cuando entre las hojas aparece aire o líquido, se separan y puede apreciarse la
existencia individualizada de cada hoja. La pleura tiene dos funciones: a)
mantener en contacto el pulmón con la pared torácica, de forma que sus
movimientos vayan al unísono, y actuar como lubricante permitiendo que las
hojas resbalen entre sí y no haya mucha fricción en un órgano en continuo
movimiento. La presencia de esa pequeña cantidad de líquido favorece de forma
extraordinaria la adherencia. La presión en la cavidad pleural es negativa, y
puede mantenerse gracias a los capilares linfáticos que drenan el líquido y
generan con su aspiración una presión negativa. La entrada de aire a la cavidad
pleural elimina la presión negativa, provocando el colapso del pulmón y
limitando de forma importante la respiración.
Vascularización
e inervación de los pulmones
El aporte de sangre a los pulmones es
tan importante para la respiración como la entrada de aire al espacio alveolar.
La circulación pulmonar dispone de una extensa red de capilares (300 millones)
que rodean cada uno de los alvéolos. La superficie total de este lecho capilar
es de unos 70 m2, lo que permite una estrecha correlación entre las superficies
alveolares y endoteliales. De esta forma se garantiza una correcta difusión de
los gases respiratorios.
Hasta las vías respiratorias llegan
fibras procedente del sistema nervioso autónomo que inervan las fibras
musculares lisas de esta zona.
Funciones del
sistema de conducción
En las cavidades nasales la presencia
de los cornetes da lugar a un incremento de superficie recubierta por un
epitelio columnar ciliado y con gran cantidad de células mucosas. Además
también hay pelos o vibrisas y una densa red de capilares a nivel de la submucosa.
Estas características estructurales, permite que el aire al penetrar en las
fosas nasales, desarrolle, por lo tortuoso de su recorrido, un flujo turbulento
que golpea contra las paredes, permitiendo así las funciones siguientes:
·
Filtrado
del aire inspirado, eliminando las partículas en suspensión que tengan un
diámetro superior a las 4-6 micras.
·
Calentamiento
del aire, por contacto con el flujo sanguíneo, pudiendo elevarse la temperatura
del aire de 2 a 3ºC.
·
Humidificación
del aire, el recorrido por las vías aéreas altas produce una saturación de
vapor de agua (100%).
·
Protección,
ya que la presencia de terminaciones nerviosas sensoriales del nervio trigémino
detectan la presencia de irritantes y produce el reflejo del estornudo.
La faringe es un conducto que conecta
la cavidad nasal con la laringe, y también la cavidad oral con el esófago y la
cavidad nasal con el oído medio. La faringe es una encrucijada de paso tanto
para el aparato respiratorio como para el digestivo, durante la deglución el
alimento procedente de la cavidad bucal es desviado de la laringe por una
lengüeta cartilaginosa, la epiglotis. La laringe es una estructura
cartilaginosa tapizada también por una mucosa ciliada que ayuda a limpiar,
humidificar y calentar el aire. Por su estructura y posición protege de la
entrada de sólidos o líquidos al aparato respiratorio y en ella se realiza la
fonación o formación de sonidos.
La pared bronquiolar está formada por
un epitelio cúbico simple sin cilios en los bronquiolos terminales, fibras
musculares lisas y fibras elásticas. Las fibras musculares lisas están
inervadas por el parasimpático (vago) que produce broncoconstricción. Existen
terminaciones sensoriales que son estimuladas por la presencia de irritantes o
por estiramiento de las paredes bronquiolares. En el epitelio hay células
mucosas que, junto con las glándulas submucosas, producen un fluído mucoso que
ayuda en el proceso de la humidificación y limpieza del aire inspirado. Este
sistema se denomina sistema escalador muco-ciliar, ya que la cubierta de mucus
es arrastrada por los cilios de las células epiteliales empujándole hacia la
faringe a una velocidad media de 2 cm/min.
Barrera
hemato-gaseosa
La barrera entre el gas situado en el
interior del alvéolo y la sangre en la densa red capilar que tapiza los
alvéolos, barrera hemato-gaseosa o membrana alvéolo-capilar, es de
aproximadamente 0,5 μ.
Los elementos que conforman esta
barrera de separación son:
·
La
capa de agua que tapiza el alvéolo en su interior.
·
El
epitelio alveolar con su membrana basal.
·
El
líquido intersticial.
·
El
endotelio capilar con su membrana basal
El epitelio alveolar está formado por
células de dos tipos:
·
Células
alveolares o neumocitos Tipo I.
·
Células
alveolares o neumocitos Tipo II.
Las células alveolares Tipo I son las más
abundantes (95%) y son células epiteliales planas o escamosas; las de Tipo II
son células cúbicas más grandes, con microvellosidades en su superficie apical,
su metabolismo es mucho más activo, sintetizan surfactante que acumulan en
cuerpos lamelares y secretan a la capa líquida que baña los alvéolos. El
surfactante es una sustancia tensoactiva, mezcla de fosfolípidos,
principalmente dipalmitoilfosfatidilcolina (lecitina), proteínas e iones, que
reduce la tensión superficial entre aire respirado y sangre, disminuye el
trabajo respiratorio y proporciona estabilidad a los alvéolos impidiendo su
colapso. También se encuentran macrófagos alveolares que recorren la superficie
alveolar y fagocitan las partículas extrañas que alcanzan el epitelio alveolar,
constituyendo la última barrera defensiva.
1
INTERCAMBIO GASEOSO
El intercambio gaseoso se lleva a
cabo realmente a través de las paredes alveolares. El aire entra y sale de los
pulmones como resultado de cambios en la presión pulmonar que, a su vez,
resultan de cambios en el tamaño de la cavidad torácica.
a) el
aire entra a través de la nariz o de la boca y pasa a la faringe, entra en la
laringe y sigue hacia abajo por la tráquea, bronquios y bronquiolos hasta los
alvéolos
b) de los
pulmones. Los alvéolos, de los que hay aproximadamente 300 millones en un par
de pulmones, son los sitios de intercambio gaseoso.
c) El
oxígeno y el dióxido de carbono difunden a través de la pared de los alvéolos y
de los capilares sanguíneos.
Desde
las cavidades nasales, el aire pasa a la faringe y desde allí a la laringe que
contiene las cuerdas vocales y está situada en la parte superior y anterior del
cuello. El aire que pasa a través de las cuerdas vocales al espirar las hace
vibrar y esto causa los sonidos del habla.
Desde la
laringe, el aire inspirado pasa a través de la tráquea, un tubo membranoso
largo también revestido de células epiteliales ciliadas.
La
tráquea desemboca en los bronquios, que se subdividen en pasajes aéreos cada
vez más pequeños llamados bronquiolos.
Los bronquios
y los bronquiolos están rodeados por capas delgadas de músculo liso. La
contracción y relajación de este músculo, que se halla bajo control del sistema
nervioso autónomo ajustan el flujo de aire según las demandas metabólicas.
Los
cilios de la tráquea, bronquios y bronquiolos baten continuamente, empujando el
moco y las partículas extrañas embebidas en él hacia la faringe, desde donde
generalmente son tragados.
El
intercambio real de gases ocurre por difusión -como consecuencia de diferentes
presiones parciales de oxígeno y dióxido de carbono- en pequeños sacos aéreos,
los alvéolos, rodeados por capilares. El endotelio de los capilares y las
células epiteliales planas de los alvéolos constituyen la barrera de difusión
entre el aire de un alvéolo y la sangre de sus capilares.
1.1
EL INTERCAMBIO DE LOS GASES POR DIFUSIÓN
dióxido
de carbono en el alvéolo y el capilar alveolar. Las cifras indican las
presiones medidas en milímetros de mercurio.
Los
pulmones están cubiertos por una membrana delgada conocida como pleura, que
también reviste la cavidad torácica. La pleura secreta una pequeña cantidad de
fluido que lubrica las superficies, de modo que éstas resbalan unas sobre otras
cuando los pulmones se expanden y se contraen.
1.1
MECANISMO DE LA RESPIRACIÓN
Los
cambios en el volumen de la cavidad torácica son los responsables de la
variación en la presión de los pulmones.
Inhalamos
contrayendo el diafragma en forma de cúpula, que aplana y alarga la cavidad
torácica, y contrayendo los músculos intercostales, que empujan la caja
torácica hacia arriba y hacia afuera. Estos movimientos agrandan la cavidad
torácica; dentro de ella disminuye la presión y el aire entra a los pulmones.
El aire es forzado a salir de los pulmones cuando los músculos se relajan y el
sistema vuelva a su equilibrio, reduciéndose el volumen de la cavidad torácica.
El
sentido del flujo aéreo en las vías respiratorias depende de la diferencia de
presión entre el alvéolo y la atmósfera. Cuando la presión alveolar es mayor
que la presión atmosférica, el aire sale y se produce la espiración. Cuando la
presión alveolar es menor que la atmosférica, el aire fluye hacia adentro y
ocurre la inspiración. Este proceso cíclico, que es la base de la ventilación,
se halla bajo control del sistema nervioso autónomo.
1.2
TRANSPORTE E INTERCAMBIO DE GASES
El
oxígeno es relativamente insoluble en el plasma sanguíneo. En animales que no
dependen de su sangre para transportar oxígeno a cada célula, ya que poseen un
sistema respiratorio traqueolar, esta baja solubilidad tiene pocas
consecuencias. En otros animales, sería una limitación grave si no fuese por la
presencia de proteínas especiales transportadoras de oxígeno -los pigmentos
respiratorios-, que elevan la capacidad de transporte de oxígeno de la sangre.
En los
vertebrados, y en muchos invertebrados el pigmento respiratorio es la
hemoglobina, que está empaquetada dentro de los glóbulos rojos. En los moluscos
y los artrópodos, la hemocianina, que contiene cobre en lugar de hierro, es el
pigmento respiratorio más común. Se conocen otros pigmentos respiratorios;
todos son una combinación de una unidad que contiene un ion metálico y una
proteína.
La hemoglobina
tiene cuatro subunidades, cada una de las cuales puede combinarse con una
molécula de oxígeno. La adición de cada molécula de oxígeno incrementa la
afinidad de la molécula por la siguiente molécula de oxígeno. Recíprocamente,
la pérdida de cada molécula de oxígeno facilita la pérdida de la molécula
siguiente.
1.3
LA CURVA DE ASOCIACIÓN-DISOCIACIÓN OXÍGENO-HEMOGLOBINA
Esta
curva representa valores de porcentaje de saturación para la hemoglobina humana
de un adulto normal a distintas presiones parciales de oxígeno, a 38° C y a pH
normal. Cuando la presión parcial de oxígeno se eleva, la hemoglobina incorpora
oxígeno. Cuando la presión de oxígeno alcanza 100 mm Hg, que es la presión
presente habitualmente en el pulmón humano, la hemoglobina se satura casi completamente
con oxígeno. Cuando la PO2 cae, el oxígeno se disocia de la hemoglobina. Por lo
tanto, cuando la sangre portadora de oxígeno alcanza los capilares, donde la
presión es sólo de 40 mm Hg o menos, libera parte de su oxígeno
(aproximadamente un 30 %) en los tejidos.
El
dióxido de carbono es más soluble que el oxígeno en la sangre y viaja, en
parte, disuelto en el plasma; en parte, unido a los grupos amino de las
moléculas de hemoglobina y, en mayor proporción, como ion bicarbonato (HCO3).
Una vez que se ha liberado en el plasma, el dióxido de carbono difunde a los
alvéolos y fluye del pulmón con el aire exhalado.
La mioglobina es un pigmento
respiratorio que se encuentra en el músculo esquelético. Estructuralmente, se
asemeja a una sola subunidad de la molécula de hemoglobina. La afinidad de la
mioglobina por el oxígeno es mayor que la de la hemoglobina, y por eso toma
oxígeno de la hemoglobina. Sin embargo, durante un ejercicio intenso, cuando
las células musculares utilizan el oxígeno rápidamente y la presión parcial de
oxígeno en las células del músculo cae a cero, la mioglobina libera su oxígeno.
De esta forma, la mioglobina suministra una reserva adicional de oxígeno a los
músculos activos.
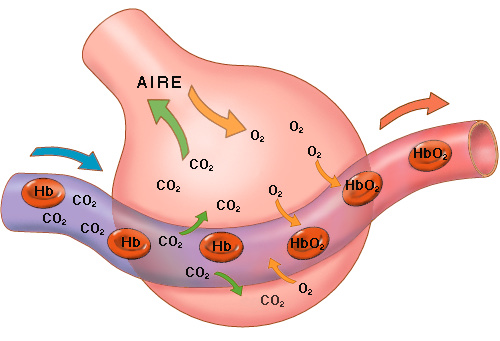
El O2 contenido en los
alvéolos (aire alveolar) pasa de éstos a la sangre y de ésta a las células. En
realidad, el intercambio de gases se realiza a dos niveles: primeramente, como
acabamos de mencionar, a nivel alveolar, lo que constituye la respiración
externa; posteriormente, a nivel celular, entre sangre, líquido, intersticial y
célula. Este intercambio es la respiración interna o tisular.
Para comprender el intercambio gaseoso,
tanto a nivel alveolar como tisular, es menester referirse a la composición del
aire ambiental, a la presión parcial de sus componentes en los alvéolos, en la
sangre y en el espacio intersticial que separa los alvéolos de los vasos
sanguíneos pulmonares. La composición del aire que queda en el espacio muerto
anatómico al final de la inspiración, es obviamente la misma que la del aire ambiental.
Su contenido en CO2 es mínimo y, por lo tanto, sin importancia.
El CO2 del aire espirado proviene, por consiguiente, de los
alvéolos, que lo han recibido a su vez de la sangre. La concentración del CO2 del
aire alveolar es, por el contrario, relativamente alta, alrededor de 6%. E1
aire espirado contiene, sin embargo, sólo 4% de CO2, debido a que es
una mezcla del aire atmosférico con el aire alveolar, que ocupa el espacio
muerto anatómico«. Esto permite calcular el volumen de este espacio muerto, a
base del siguiente razonamiento: la concentración de CO2 en el
aire espirado es de 4% y la del aire alveolar de 6%. Esto indica que una
tercera parte del CO2 alveolar se ha diluido en el aire del
espacio muerto anatómico cuyo volumen debe ser, entonces, alrededor de 150 ml.
La ventilación pulmonar aumenta paralelamente con la frecuencia respiratoria
sólo hasta cierto límite.
Con la primera inspiración después del
parto, el aire penetra del espacio muerto anatómico a los alvéolos,
expandiéndolos por primera vez y se convierte en aire alveolar. El volumen de
aire alveolar que se expulsa durante la espiración, se mezcla con el aire del
espacio muerto y constituye el aire espirado. La cantidad de
aire atmosférico que llega a penetrar a los alvéolos por cada inspiración
es, por consiguiente, bastante pequeña (350 ml) en comparación con la cantidad
total de aire alveolar al final de una espiración tranquila. La ventilación
alveolar utiliza, por lo tanto, sólo parcialmente los gases del aire ambiental.
Este hecho explica la composición diferente del aire ambiental y alveolar.
El O2 pasa de los
alvéolos a los capilares pulmonares y simultáneamente el CO2 de
éstos a los alvéolos. Durante este intercambio la concentración del O2 del aire
alveolar disminuye y se eleva la del CO2, para retornar a sus
niveles primitivos en la próxima inspiración. Gracias a la ventilación
pulmonar, la composición del aire alveolar se mantiene prácticamente constante.
Los aires inspirado, espirado y
alveolar tienen la siguiente composición:

El aire alveolar es
una mezcla de gases, en que cada uno de los componentes ejerce separadamente su
presión parcial. La suma de las presiones parciales es igual a la presión total
del aire alveolar. Existe, además, entre las presiones parciales de los gases
del aire inspirado y del aire alveolar otra diferencia que se debe a la
saturación del aire alveolar con vapor de agua. La presión parcial del vapor de
agua, a una temperatura de 38º C y a nivel del mar (presión atmosférica igual
a 760 mm de Hg), es alrededor de 47 mm de Hg. Por consiguiente, la presión
conjunta del O2, CO2 y N2 en el aire
alveolar y en el espirado, es inferior en aproximadamente 47 mm de Hg a la que
tienen en el aire inspirado (atmosférico). La saturación con vapor de agua disminuye
también en forma proporcional las respectivas presiones parciales.
Conociendo la
composición porcentual de una mezcla de gases y la presión total de la misma,
es fácil calcular la presión parcial que corresponde a cada uno de dichos
gases. Aplicando lo anterior a los gases respiratorios, tenemos que a nivel del
mar la presión total del aire alveolar será de 713 mm de Hg. De ésta, 47 mm de
Hg corresponde a la presión de vapor de agua así que la presión total del resto
de los gases tendrá sólo un valor de 713 mm de Hg. De esta presión corresponde
un 14% (100 mm de Hg) al O2, un 5.6% (40 mm de Hg) al CO2 y
el restante 80.4% (573 mm de Hg) al N2.
Expresado en otra
forma:

2
PRESIONES RESPIRATORIAS
Por convenio en el aparato
respiratorio las presiones se miden tomando como referencia la presión
atmosférica. Una presión será negativa cuando sea menor de 760 mmHg y positiva
si es mayor. Durante la inhalación normal la presión dentro de los pulmones
presión intralveolar, es cerca de -2 cm de agua.
La presión, generada por la
fuerza de contracción de los músculos inspiratorios tiene que compensar:
La
fuerza de retroceso elástica del pulmón
La disposición de los
alveolos y la presencia de elastina en su estructura les confieren propiedades
semejantes a las de un resorte regido por la ley de Hooke:
Para mantener un elemento
elástico como el alveolo con un determinado volumen se requiere una presión que
compense la fuerza elástica. Esto se estudia representando la relación entre
presión y volumen.
La tensión superficial de la interface aire
liquido
En 1929 von Neergaard
descubrió que si se inflaba un pulmón con líquido la presión que se necesitaba
era mucho menor que cuando se utilizaba aire. Dedujo que esto se debía a que el
líquido suprimía la interface aire líquido y eliminaba la fuerza de tensión
superficial. Cuando el alvéolo se expande con aire se genera una fuerza de
tensión superficial que se opone al desplazamiento y que debe ser compensada
por la presión de acuerdo con la ley de Laplace.
Sin embargo el pulmón tiene
un comportamiento peculiar. En primer lugar la fuerza de tensión superficial es
menor que la que se desarrolla en una interface aire plasma. Esto se explica
por la existencia, en los alvéolos, de unas células, los neumocitos tipo II,
que secretan un agente tensioactivo el surfactante que modifica la tensión
interfacial: a mayor concentración de surfactante, menor es la tensión
superficial. En segundo lugar de la ley de Laplace se deduce que si la tensión
superficial es constante, la presión de equilibrio tiene que ser mayor en los
alvéolos pequeños que en los grandes. Como los alvéolos están intercomunicados,
los más pequeños se vaciarían en los mayores y un sistema con alvéolos de
distinto tamaño sería inestable.
Esto no sucede en la
realidad y se debe justamente a la presencia del surfactante alveolar. La masa
o cantidad de surfactante permanece constante en el alvéolo, mas no su
proporción por unidad de superficie alveolar; es decir, su concentración
superficial cambia con el volumen. Al expandirse el alvéolo durante una
inspiración su área se incrementa, pero al permanecer constante la masa del
surfactante, la concentración superficial o cantidad del mismo por unidad de
área alveolar se vuelve más pequeña; como resultado, incrementa la tensión
superficial. Lo contrario ocurre cuando el alvéolo se contrae: disminuye su
área y aumenta la concentración superficial de surfactante, con lo cual se
reduce la tensión superficial.
En particular en los
alveolos pequeños la tensión superficial puede ser hasta diez veces menor que
en los mayores. De esta manera, al cambiar el numerador y el denominador en la
relación de Laplace se explica que puedan coexistir alvéolos de distinto tamaño
con la misma presión intraalveolar. Este mismo fenómeno también coopera en la
histéresis que presenta el pulmón, al existir una diferencia entre la
distensibilidad pulmonar durante la inflación y deflación ya
que la histéresis es mucho menor cuando el pulmón se rellena con líquido en
lugar de con aire.
La fuerza elástica y la
tensión superficial se analizan, en las pruebas funcionales respiratorias,
mediante la adaptabilidad pulmonar (llamada también distensibilidad o
comparanza) que es el cambio de volumen que produce un cambio de una unidad de
presión y cuyo valor normal es de unos 0,2 litros de aire por cada cm de agua
de presión.
LA RESISTENCIA AL FLUJO
Durante el movimiento
pulmonar (condiciones dinámicas) la presión debe compensar también la
resistencia al flujo. En gran parte de las vías aéreas el flujo se puede
considerar laminar y viene regido por la ley de Poiseuille:
Es el factor más
importante, porque es el que puede cambiar en el organismo y porque interviene
en su cuarta potencia el calibre de los bronquios, de ahí los efectos
dramáticos que puede ca usar la bronquioconstricción. En las grandes vías
respiratorias como la tráquea y los grandes bronquios el flujo puede ser
turbulento y entonces la presión se relaciona con el flujo y con el cuadrado
del flujo y la resistencia depende de la densidad más que de la viscosidad.
Esto es importante cuando se respira aire a presión (como en el buceo) ya que,
en los gases, la densidad es proporcional a la presión. En las pruebas
funcionales respiratorias la resistencia de las vías aéreas se estudia mediante
las curvas de flujo-volumen.
2.1
VOLÚMENES Y CAPACIDADES
PULMONARES ESTÁTICOS
Los volúmenes pulmonares
estáticos son un reflejo de las propiedades elásticas de los pulmones y de la
caja torácica. La capacidad vital (VC = Vital Capacity, ó "VC lenta")
es la combinación del volumen tidal o de corriente, del volumen de reserva
inspiratoria y del volumen de reserva espiratoria. Representa el volumen total
de aire que se puede inspirar después de una máxima expiracion.2 Dado que la VC
disminuye a medida que las enfermedades restrictivas empeoran, ésta junto con
la capacidad de transferencia de pueden ser utilizados como parámetros básicos
al efectuar un seguimiento de la evolución de una enfermedad pulmonar
restrictiva y por tanto de su respuesta al tratamiento.
La capacidad vital forzada
(FVC = Torced Vital Capacity), es una maniobra parecida a la anterior a
excepción de que se requiere de una espiración forzada (rápida) máxima, por lo
general se mide junto a los flujos espiratorios máximos en la espirómetro
simple.
La VC (lenta) puede ser
considerablemente mayor que la FVC en pacientes con obstrucción de la vía
aérea. Durante la maniobra de FVC, las vías aéreas terminales pueden cerrarse
de forma prematura (es decir, antes de que se alcance el volumen residual
verdadero), atrapando gas en sus porciones distales y evitando que éste sea
medido por el espirómetro.
La capacidad pulmonar total
(TLC = Total Lung Capacity) es el volumen de aire que permanece dentro de los
pulmones al final de una inspiración máxima
La capacidad residual funcional
(FRC = Functional Residual Capacity) es el volumen de aire contenido en los
pulmones al final de una espiración normal, cuando todos los músculos
respiratorios están relajados. Fisiológicamente, es el volumen pulmonar de
mayor importancia, dada su proximidad al rango normal del volumen corriente. Al
nivel de la FRC, las fuerzas de retracción elástica de la pared torácica, que
tienden a aumentar el volumen pulmonar, se hallan en equilibrio con las del
parénquima pulmonar, que tienden a reducirla.
En condiciones normales,
estas fuerzas son iguales y de sentido opuesto, aproximadamente el 40% de la
TLC. Los cambios de estas propiedades elásticas modifican la FRC. La pérdida de
retracción elástica del pulmón en el enfisema aumenta el
valor de la FRC. Por el contrario, el aumento de la rigidez pulmonar que se
asocia al edema pulmonar, la fibrosis intersticial, y otras enfermedades
restrictivas provoca disminución de la FRC. La cifoscoliosis disminuye la FRC y
otros volúmenes pulmonares, debido a que la pared torácica rígida y no
distensible restringe la expansión pulmonar. La diferencia entre la TLC y la
FRC es la capacidad inspiratoria.
2.2
VOLÚMENES PULMONARES Y FLUJOS
AÉREOS DINÁMICOS
Los volúmenes pulmonares
dinámicos reflejan el estado de las vías aéreas. El espirograma proporciona una
gráfica de volumen contra tiempo, obtenida en un espirómetro de campana o
electrónico, mientras el enfermo realiza una maniobra de FVC. El VEF1 (o FEV1
por sus siglas en inglés Forced Expiratory Volume in the first second) es el
volumen de aire eliminado durante el primer segundo de espiración forzada,
después de una inspiración máxima; en condiciones normales, su valor es mayor
al 75 % de la VC, por lo que a menudo se expresa en forma de porcentaje de la
capacidad vital forzada (FEV1% FVC).
El índice de Tiffenau es la
relación entre la FEV1 y la CV:
·
en
individuos normales, suele oscilar
alrededor del 80%;
·
en
pacientes con enfermedades obstructivas (como asma, EPOC o enfisema), suele
representar el 30-40%, dado que la FEV1 disminuye mucho más que la FVC;
·
en
pacientes con enfermedades restrictivas, suele obtenerse un valor normal (como
en la enfermedad de Duchenne) o mayor (como en la fibrosis pulmonar), porque la
FEV1 y la FVC disminuyen de forma paralela.
El flujo espiratorio forzado
medio (FEF25-75 %) durante la fase media (del 25% al 75 %) de la maniobra de
FVC es la pendiente de la línea que corta el trazado espirográfico al 25% y al
75 % de la VC. El FEF25-75 % depende menos del esfuerzo realizado que el FEV1
y, por lo tanto, constituye un indicador más precoz de obstrucción de las vías
aéreas.
En una curva de flujo
volumen normal, la porción inspiratoria de la curva es simétrica y convexa. La
porción respiratoria es lineal. Los flujos se miden a menudo en el punto medio
de la VC. El MIF50% es > MEF50% VC debido a la compresión dinámica de las
vías aéreas.
En ocasiones se utiliza el
flujo respiratorio máximo para estimar el grado de obstrucción de la vía aérea,
pero depende mucho del esfuerzo realizado por el paciente. Los flujos
espiratorios medidos por encima del 50% de la VC, es decir, cercanos al RV son
indicadores sensibles del estado de las vías aéreas del pequeño calibre.
En una enfermedad
restrictiva, por ejemplo sarcoidosis ó cifoscoliosis. La curva es más estrecha
a causa de la reducción de los volúmenes pulmonares
Durante una maniobra de
espiración forzada, la presión intratorácica positiva determina que las vías
aéreas se vayan estrechando de modo progresivo. Esta compresión dinámica de las
vías aéreas limita las velocidades máximas de flujo respiratorio que pueden
alcanzarse. Durante la maniobra de inspiración se produce el efecto opuesto, ya
que la presión intratorácica negativa tiende a mantener al máximo el calibre de
las vías aéreas. Debido a estas variaciones de diámetro de las vías aéreas, en
la mayor parte del ciclo respiratorio las velocidades de flujo aéreo son mucho
mayores durante la inspiración que durante la espiración. La ventilación
voluntaria máxima (MVV = Máxima Voluntaria Ventilación) se calcula indicando al
enfermo que respire durante 15 segundos a volumen y frecuencia respiratoria
máximos (la cantidad de aire espirado se expresa en lt/min.). En general, el
valor de la MVV es paralelo al del FEV1, y puede aplicarse una fórmula simple
para comprobar la uniformidad interna de la prueba y valorar el grado de
cooperación de enfermo. Es posible predecir la MVV a partir
del espirograma, multiplicando el FEV1 (en lt) x 35 o 40, según los autores.
Esta fórmula sirve tanto para los individuos sanos como para los enfermos con
trastornos respiratorios obstructivos y restrictivos.
Si se observa una MVV muy
baja en un usuario que parece cooperar de forma activa, hay que pensar en una
debilidad neuromuscular. Exceptuando los casos de enfermedad neuromuscular muy
avanzada, la mayoría de los usuarios son capaces de efectuar un esfuerzo
respiratorio aislado como un FVC. La MVV requiere un esfuerzo mucho mayor, y su
alteración demuestra la existencia de músculos respiratorios débiles y
fatigables. La MVV disminuye progresivamente cuando existe un aumento de la
debilidad de los músculos respiratorios; junto con las presiones inspiratoria y
espiratoria máximas, la MVV es en ocasiones, la única prueba funcional
respiratoria anómala en ciertos individuos con una enfermedad neuromuscular
relativamente grave.
La MVV es importante
también en la valoración del riesgo quirúrgico, pues refleja la gravedad de la
obstrucción de las vías aéreas y también las reservas respiratorias, la fuerza
muscular y el grado de motivación del usuario. Volúmenes pulmonares, valor de
volúmenes, capacidad total, capacidad residual funcional, capacidad reserva
respiratoria.
3
COLAPSO PULMONAR
El colapso pulmonar ocurre
cuando el aire escapa del pulmón. El aire luego llena el espacio por fuera del
pulmón, entre éste y la pared torácica. Esta acumulación de aire ejerce presión
sobre el pulmón, así que éste no se puede expandir tanto como lo hace
normalmente cuando usted inspira.
El término médico para esta
afección es neumotórax.
Causas
El colapso pulmonar puede
ser causado por una lesión al pulmón. Las lesiones pueden abarcar herida por
arma de fuego o cuchillo en el tórax, fractura de una costilla o ciertos
procedimientos médicos.
En algunos casos, un
colapso pulmonar es causado por ampollas de aire (vesículas) que se rompen,
enviando aire hacia el espacio que rodea el pulmón. Esto puede resultar de los
cambios de presión del aire como sucede al bucear o viajar a grandes alturas.
Las personas altas y
delgadas y los fumadores tienen mayor probabilidad de sufrir un colapso
pulmonar.
Las neumopatías también
pueden incrementar la posibilidad de sufrir un colapso pulmonar y son, entre
otras:
·
Asma
·
EPOC
·
Fibrosis
quística
·
Tuberculosis
·
Tos
ferina
En algunos casos, un
colapso pulmonar ocurre sin ninguna causa. Esto se denomina neumotórax
espontáneo o atelectasia pulmonar.
La ventilación mecánica es
una estrategia terapéutica que consiste en remplazar o asistir mecánicamente la
ventilación pulmonar espontánea cuando ésta es inexistente o ineficaz para la
vida. Para llevar a cabo la ventilación mecánica se puede recurrir o bien a una
máquina (ventilador mecánico) o bien a una persona bombeando el aire
manualmente mediante la compresión de una bolsa o fuelle de aire.
Se llama ventilación
pulmonar al intercambio de gases entre los pulmones y la atmósfera. Tiene como
fin permitir la oxigenación de la sangre (captación de oxígeno) y la
eliminación de dióxido de carbono.
En la ventilación
espontánea, durante la inspiración, un individuo genera presiones
intratorácicas negativas al aumentar el volumen torácico gracias a la
musculatura respiratoria (principalmente el diafragma). La presión en el
interior del tórax se hace menor que la atmosférica, generando así un gradiente
de presiones que provoca la entrada de aire a los pulmones para equilibrar esa
diferencia. La espiración (salida de aire) normalmente es un proceso pasivo.
Durante la ventilación
espontánea se introduce y expulsa un volumen regular de aire llamado volumen
tidal, de aproximadamente ½ litro, a una frecuencia respiratoria determinada
(12 – 20 respiraciones por minuto).
3.1
MECANISMOS
QUE CAUSAN UN COLAPSO PULMONAR
Son dos
mecanismos
•
La presión
intrapleural negativa ayuda a mantener los pulmones distendidos.
•
La sustancia tensioactiva o surfactante disminuye la
tensión superficial de los liquídos que rodean a los alvéolos. La sustancia
tensioactiva es una mezcla bifásica de proteínas fosfolípidos e iones ;
«dipalmitoil- lecitina» (apoproteiina surfactante e iones de calcio). Previene
el edema pulmonar, es producido por el epitelio alveolar las células tipo II.
3.2
MECANISMOS QUE LLEVAN A UN
COPALSO PULMONAR
Depende de dos
factores
•
Las fibras elásticas del
parénquima pulmonar.
•
La tensión superficial de
los líquidos que recubren a los alvéolos.
dióxido
de carbono en el alvéolo y el capilar alveolar. Las cifras indican las
presiones medidas en milímetros de mercurio.
Los
pulmones están cubiertos por una membrana delgada conocida como pleura, que
también reviste la cavidad torácica. La pleura secreta una pequeña cantidad de
fluido que lubrica las superficies, de modo que éstas resbalan unas sobre otras
cuando los pulmones se expanden y se contraen.
1.1
MECANISMO DE LA RESPIRACIÓN
Los
cambios en el volumen de la cavidad torácica son los responsables de la
variación en la presión de los pulmones.
Inhalamos
contrayendo el diafragma en forma de cúpula, que aplana y alarga la cavidad
torácica, y contrayendo los músculos intercostales, que empujan la caja
torácica hacia arriba y hacia afuera. Estos movimientos agrandan la cavidad
torácica; dentro de ella disminuye la presión y el aire entra a los pulmones.
El aire es forzado a salir de los pulmones cuando los músculos se relajan y el
sistema vuelva a su equilibrio, reduciéndose el volumen de la cavidad torácica.
El
sentido del flujo aéreo en las vías respiratorias depende de la diferencia de
presión entre el alvéolo y la atmósfera. Cuando la presión alveolar es mayor
que la presión atmosférica, el aire sale y se produce la espiración. Cuando la
presión alveolar es menor que la atmosférica, el aire fluye hacia adentro y
ocurre la inspiración. Este proceso cíclico, que es la base de la ventilación,
se halla bajo control del sistema nervioso autónomo.
1.2
TRANSPORTE E INTERCAMBIO DE GASES
El
oxígeno es relativamente insoluble en el plasma sanguíneo. En animales que no
dependen de su sangre para transportar oxígeno a cada célula, ya que poseen un
sistema respiratorio traqueolar, esta baja solubilidad tiene pocas
consecuencias. En otros animales, sería una limitación grave si no fuese por la
presencia de proteínas especiales transportadoras de oxígeno -los pigmentos
respiratorios-, que elevan la capacidad de transporte de oxígeno de la sangre.
En los
vertebrados, y en muchos invertebrados el pigmento respiratorio es la
hemoglobina, que está empaquetada dentro de los glóbulos rojos. En los moluscos
y los artrópodos, la hemocianina, que contiene cobre en lugar de hierro, es el
pigmento respiratorio más común. Se conocen otros pigmentos respiratorios;
todos son una combinación de una unidad que contiene un ion metálico y una
proteína.
La hemoglobina
tiene cuatro subunidades, cada una de las cuales puede combinarse con una
molécula de oxígeno. La adición de cada molécula de oxígeno incrementa la
afinidad de la molécula por la siguiente molécula de oxígeno. Recíprocamente,
la pérdida de cada molécula de oxígeno facilita la pérdida de la molécula
siguiente.
1.3
LA CURVA DE ASOCIACIÓN-DISOCIACIÓN OXÍGENO-HEMOGLOBINA
Esta
curva representa valores de porcentaje de saturación para la hemoglobina humana
de un adulto normal a distintas presiones parciales de oxígeno, a 38° C y a pH
normal. Cuando la presión parcial de oxígeno se eleva, la hemoglobina incorpora
oxígeno. Cuando la presión de oxígeno alcanza 100 mm Hg, que es la presión
presente habitualmente en el pulmón humano, la hemoglobina se satura casi completamente
con oxígeno. Cuando la PO2 cae, el oxígeno se disocia de la hemoglobina. Por lo
tanto, cuando la sangre portadora de oxígeno alcanza los capilares, donde la
presión es sólo de 40 mm Hg o menos, libera parte de su oxígeno
(aproximadamente un 30 %) en los tejidos.
El
dióxido de carbono es más soluble que el oxígeno en la sangre y viaja, en
parte, disuelto en el plasma; en parte, unido a los grupos amino de las
moléculas de hemoglobina y, en mayor proporción, como ion bicarbonato (HCO3).
Una vez que se ha liberado en el plasma, el dióxido de carbono difunde a los
alvéolos y fluye del pulmón con el aire exhalado.
La mioglobina es un pigmento
respiratorio que se encuentra en el músculo esquelético. Estructuralmente, se
asemeja a una sola subunidad de la molécula de hemoglobina. La afinidad de la
mioglobina por el oxígeno es mayor que la de la hemoglobina, y por eso toma
oxígeno de la hemoglobina. Sin embargo, durante un ejercicio intenso, cuando
las células musculares utilizan el oxígeno rápidamente y la presión parcial de
oxígeno en las células del músculo cae a cero, la mioglobina libera su oxígeno.
De esta forma, la mioglobina suministra una reserva adicional de oxígeno a los
músculos activos.
El O2 contenido en los
alvéolos (aire alveolar) pasa de éstos a la sangre y de ésta a las células. En
realidad, el intercambio de gases se realiza a dos niveles: primeramente, como
acabamos de mencionar, a nivel alveolar, lo que constituye la respiración
externa; posteriormente, a nivel celular, entre sangre, líquido, intersticial y
célula. Este intercambio es la respiración interna o tisular.
Para comprender el intercambio gaseoso,
tanto a nivel alveolar como tisular, es menester referirse a la composición del
aire ambiental, a la presión parcial de sus componentes en los alvéolos, en la
sangre y en el espacio intersticial que separa los alvéolos de los vasos
sanguíneos pulmonares. La composición del aire que queda en el espacio muerto
anatómico al final de la inspiración, es obviamente la misma que la del aire ambiental.
Su contenido en CO2 es mínimo y, por lo tanto, sin importancia.
El CO2 del aire espirado proviene, por consiguiente, de los
alvéolos, que lo han recibido a su vez de la sangre. La concentración del CO2 del
aire alveolar es, por el contrario, relativamente alta, alrededor de 6%. E1
aire espirado contiene, sin embargo, sólo 4% de CO2, debido a que es
una mezcla del aire atmosférico con el aire alveolar, que ocupa el espacio
muerto anatómico«. Esto permite calcular el volumen de este espacio muerto, a
base del siguiente razonamiento: la concentración de CO2 en el
aire espirado es de 4% y la del aire alveolar de 6%. Esto indica que una
tercera parte del CO2 alveolar se ha diluido en el aire del
espacio muerto anatómico cuyo volumen debe ser, entonces, alrededor de 150 ml.
La ventilación pulmonar aumenta paralelamente con la frecuencia respiratoria
sólo hasta cierto límite.
Con la primera inspiración después del
parto, el aire penetra del espacio muerto anatómico a los alvéolos,
expandiéndolos por primera vez y se convierte en aire alveolar. El volumen de
aire alveolar que se expulsa durante la espiración, se mezcla con el aire del
espacio muerto y constituye el aire espirado. La cantidad de
aire atmosférico que llega a penetrar a los alvéolos por cada inspiración
es, por consiguiente, bastante pequeña (350 ml) en comparación con la cantidad
total de aire alveolar al final de una espiración tranquila. La ventilación
alveolar utiliza, por lo tanto, sólo parcialmente los gases del aire ambiental.
Este hecho explica la composición diferente del aire ambiental y alveolar.
El O2 pasa de los
alvéolos a los capilares pulmonares y simultáneamente el CO2 de
éstos a los alvéolos. Durante este intercambio la concentración del O2 del aire
alveolar disminuye y se eleva la del CO2, para retornar a sus
niveles primitivos en la próxima inspiración. Gracias a la ventilación
pulmonar, la composición del aire alveolar se mantiene prácticamente constante.
Los aires inspirado, espirado y
alveolar tienen la siguiente composición:
El aire alveolar es
una mezcla de gases, en que cada uno de los componentes ejerce separadamente su
presión parcial. La suma de las presiones parciales es igual a la presión total
del aire alveolar. Existe, además, entre las presiones parciales de los gases
del aire inspirado y del aire alveolar otra diferencia que se debe a la
saturación del aire alveolar con vapor de agua. La presión parcial del vapor de
agua, a una temperatura de 38º C y a nivel del mar (presión atmosférica igual
a 760 mm de Hg), es alrededor de 47 mm de Hg. Por consiguiente, la presión
conjunta del O2, CO2 y N2 en el aire
alveolar y en el espirado, es inferior en aproximadamente 47 mm de Hg a la que
tienen en el aire inspirado (atmosférico). La saturación con vapor de agua disminuye
también en forma proporcional las respectivas presiones parciales.
Conociendo la
composición porcentual de una mezcla de gases y la presión total de la misma,
es fácil calcular la presión parcial que corresponde a cada uno de dichos
gases. Aplicando lo anterior a los gases respiratorios, tenemos que a nivel del
mar la presión total del aire alveolar será de 713 mm de Hg. De ésta, 47 mm de
Hg corresponde a la presión de vapor de agua así que la presión total del resto
de los gases tendrá sólo un valor de 713 mm de Hg. De esta presión corresponde
un 14% (100 mm de Hg) al O2, un 5.6% (40 mm de Hg) al CO2 y
el restante 80.4% (573 mm de Hg) al N2.
Expresado en otra
forma:
2
PRESIONES RESPIRATORIAS
Por convenio en el aparato
respiratorio las presiones se miden tomando como referencia la presión
atmosférica. Una presión será negativa cuando sea menor de 760 mmHg y positiva
si es mayor. Durante la inhalación normal la presión dentro de los pulmones
presión intralveolar, es cerca de -2 cm de agua.
La presión, generada por la
fuerza de contracción de los músculos inspiratorios tiene que compensar:
La
fuerza de retroceso elástica del pulmón
La disposición de los
alveolos y la presencia de elastina en su estructura les confieren propiedades
semejantes a las de un resorte regido por la ley de Hooke:
Para mantener un elemento
elástico como el alveolo con un determinado volumen se requiere una presión que
compense la fuerza elástica. Esto se estudia representando la relación entre
presión y volumen.
La tensión superficial de la interface aire
liquido
En 1929 von Neergaard
descubrió que si se inflaba un pulmón con líquido la presión que se necesitaba
era mucho menor que cuando se utilizaba aire. Dedujo que esto se debía a que el
líquido suprimía la interface aire líquido y eliminaba la fuerza de tensión
superficial. Cuando el alvéolo se expande con aire se genera una fuerza de
tensión superficial que se opone al desplazamiento y que debe ser compensada
por la presión de acuerdo con la ley de Laplace.
Sin embargo el pulmón tiene
un comportamiento peculiar. En primer lugar la fuerza de tensión superficial es
menor que la que se desarrolla en una interface aire plasma. Esto se explica
por la existencia, en los alvéolos, de unas células, los neumocitos tipo II,
que secretan un agente tensioactivo el surfactante que modifica la tensión
interfacial: a mayor concentración de surfactante, menor es la tensión
superficial. En segundo lugar de la ley de Laplace se deduce que si la tensión
superficial es constante, la presión de equilibrio tiene que ser mayor en los
alvéolos pequeños que en los grandes. Como los alvéolos están intercomunicados,
los más pequeños se vaciarían en los mayores y un sistema con alvéolos de
distinto tamaño sería inestable.
Esto no sucede en la
realidad y se debe justamente a la presencia del surfactante alveolar. La masa
o cantidad de surfactante permanece constante en el alvéolo, mas no su
proporción por unidad de superficie alveolar; es decir, su concentración
superficial cambia con el volumen. Al expandirse el alvéolo durante una
inspiración su área se incrementa, pero al permanecer constante la masa del
surfactante, la concentración superficial o cantidad del mismo por unidad de
área alveolar se vuelve más pequeña; como resultado, incrementa la tensión
superficial. Lo contrario ocurre cuando el alvéolo se contrae: disminuye su
área y aumenta la concentración superficial de surfactante, con lo cual se
reduce la tensión superficial.
En particular en los
alveolos pequeños la tensión superficial puede ser hasta diez veces menor que
en los mayores. De esta manera, al cambiar el numerador y el denominador en la
relación de Laplace se explica que puedan coexistir alvéolos de distinto tamaño
con la misma presión intraalveolar. Este mismo fenómeno también coopera en la
histéresis que presenta el pulmón, al existir una diferencia entre la
distensibilidad pulmonar durante la inflación y deflación ya
que la histéresis es mucho menor cuando el pulmón se rellena con líquido en
lugar de con aire.
La fuerza elástica y la
tensión superficial se analizan, en las pruebas funcionales respiratorias,
mediante la adaptabilidad pulmonar (llamada también distensibilidad o
comparanza) que es el cambio de volumen que produce un cambio de una unidad de
presión y cuyo valor normal es de unos 0,2 litros de aire por cada cm de agua
de presión.
LA RESISTENCIA AL FLUJO
Durante el movimiento
pulmonar (condiciones dinámicas) la presión debe compensar también la
resistencia al flujo. En gran parte de las vías aéreas el flujo se puede
considerar laminar y viene regido por la ley de Poiseuille:
Es el factor más
importante, porque es el que puede cambiar en el organismo y porque interviene
en su cuarta potencia el calibre de los bronquios, de ahí los efectos
dramáticos que puede ca usar la bronquioconstricción. En las grandes vías
respiratorias como la tráquea y los grandes bronquios el flujo puede ser
turbulento y entonces la presión se relaciona con el flujo y con el cuadrado
del flujo y la resistencia depende de la densidad más que de la viscosidad.
Esto es importante cuando se respira aire a presión (como en el buceo) ya que,
en los gases, la densidad es proporcional a la presión. En las pruebas
funcionales respiratorias la resistencia de las vías aéreas se estudia mediante
las curvas de flujo-volumen.
2.1
VOLÚMENES Y CAPACIDADES
PULMONARES ESTÁTICOS
Los volúmenes pulmonares
estáticos son un reflejo de las propiedades elásticas de los pulmones y de la
caja torácica. La capacidad vital (VC = Vital Capacity, ó "VC lenta")
es la combinación del volumen tidal o de corriente, del volumen de reserva
inspiratoria y del volumen de reserva espiratoria. Representa el volumen total
de aire que se puede inspirar después de una máxima expiracion.2 Dado que la VC
disminuye a medida que las enfermedades restrictivas empeoran, ésta junto con
la capacidad de transferencia de pueden ser utilizados como parámetros básicos
al efectuar un seguimiento de la evolución de una enfermedad pulmonar
restrictiva y por tanto de su respuesta al tratamiento.
La capacidad vital forzada
(FVC = Torced Vital Capacity), es una maniobra parecida a la anterior a
excepción de que se requiere de una espiración forzada (rápida) máxima, por lo
general se mide junto a los flujos espiratorios máximos en la espirómetro
simple.
La VC (lenta) puede ser
considerablemente mayor que la FVC en pacientes con obstrucción de la vía
aérea. Durante la maniobra de FVC, las vías aéreas terminales pueden cerrarse
de forma prematura (es decir, antes de que se alcance el volumen residual
verdadero), atrapando gas en sus porciones distales y evitando que éste sea
medido por el espirómetro.
La capacidad pulmonar total
(TLC = Total Lung Capacity) es el volumen de aire que permanece dentro de los
pulmones al final de una inspiración máxima
La capacidad residual funcional
(FRC = Functional Residual Capacity) es el volumen de aire contenido en los
pulmones al final de una espiración normal, cuando todos los músculos
respiratorios están relajados. Fisiológicamente, es el volumen pulmonar de
mayor importancia, dada su proximidad al rango normal del volumen corriente. Al
nivel de la FRC, las fuerzas de retracción elástica de la pared torácica, que
tienden a aumentar el volumen pulmonar, se hallan en equilibrio con las del
parénquima pulmonar, que tienden a reducirla.
En condiciones normales,
estas fuerzas son iguales y de sentido opuesto, aproximadamente el 40% de la
TLC. Los cambios de estas propiedades elásticas modifican la FRC. La pérdida de
retracción elástica del pulmón en el enfisema aumenta el
valor de la FRC. Por el contrario, el aumento de la rigidez pulmonar que se
asocia al edema pulmonar, la fibrosis intersticial, y otras enfermedades
restrictivas provoca disminución de la FRC. La cifoscoliosis disminuye la FRC y
otros volúmenes pulmonares, debido a que la pared torácica rígida y no
distensible restringe la expansión pulmonar. La diferencia entre la TLC y la
FRC es la capacidad inspiratoria.
2.2
VOLÚMENES PULMONARES Y FLUJOS
AÉREOS DINÁMICOS
Los volúmenes pulmonares
dinámicos reflejan el estado de las vías aéreas. El espirograma proporciona una
gráfica de volumen contra tiempo, obtenida en un espirómetro de campana o
electrónico, mientras el enfermo realiza una maniobra de FVC. El VEF1 (o FEV1
por sus siglas en inglés Forced Expiratory Volume in the first second) es el
volumen de aire eliminado durante el primer segundo de espiración forzada,
después de una inspiración máxima; en condiciones normales, su valor es mayor
al 75 % de la VC, por lo que a menudo se expresa en forma de porcentaje de la
capacidad vital forzada (FEV1% FVC).
El índice de Tiffenau es la
relación entre la FEV1 y la CV:
·
en
individuos normales, suele oscilar
alrededor del 80%;
·
en
pacientes con enfermedades obstructivas (como asma, EPOC o enfisema), suele
representar el 30-40%, dado que la FEV1 disminuye mucho más que la FVC;
·
en
pacientes con enfermedades restrictivas, suele obtenerse un valor normal (como
en la enfermedad de Duchenne) o mayor (como en la fibrosis pulmonar), porque la
FEV1 y la FVC disminuyen de forma paralela.
El flujo espiratorio forzado
medio (FEF25-75 %) durante la fase media (del 25% al 75 %) de la maniobra de
FVC es la pendiente de la línea que corta el trazado espirográfico al 25% y al
75 % de la VC. El FEF25-75 % depende menos del esfuerzo realizado que el FEV1
y, por lo tanto, constituye un indicador más precoz de obstrucción de las vías
aéreas.
En una curva de flujo
volumen normal, la porción inspiratoria de la curva es simétrica y convexa. La
porción respiratoria es lineal. Los flujos se miden a menudo en el punto medio
de la VC. El MIF50% es > MEF50% VC debido a la compresión dinámica de las
vías aéreas.
En ocasiones se utiliza el
flujo respiratorio máximo para estimar el grado de obstrucción de la vía aérea,
pero depende mucho del esfuerzo realizado por el paciente. Los flujos
espiratorios medidos por encima del 50% de la VC, es decir, cercanos al RV son
indicadores sensibles del estado de las vías aéreas del pequeño calibre.
En una enfermedad
restrictiva, por ejemplo sarcoidosis ó cifoscoliosis. La curva es más estrecha
a causa de la reducción de los volúmenes pulmonares
Durante una maniobra de
espiración forzada, la presión intratorácica positiva determina que las vías
aéreas se vayan estrechando de modo progresivo. Esta compresión dinámica de las
vías aéreas limita las velocidades máximas de flujo respiratorio que pueden
alcanzarse. Durante la maniobra de inspiración se produce el efecto opuesto, ya
que la presión intratorácica negativa tiende a mantener al máximo el calibre de
las vías aéreas. Debido a estas variaciones de diámetro de las vías aéreas, en
la mayor parte del ciclo respiratorio las velocidades de flujo aéreo son mucho
mayores durante la inspiración que durante la espiración. La ventilación
voluntaria máxima (MVV = Máxima Voluntaria Ventilación) se calcula indicando al
enfermo que respire durante 15 segundos a volumen y frecuencia respiratoria
máximos (la cantidad de aire espirado se expresa en lt/min.). En general, el
valor de la MVV es paralelo al del FEV1, y puede aplicarse una fórmula simple
para comprobar la uniformidad interna de la prueba y valorar el grado de
cooperación de enfermo. Es posible predecir la MVV a partir
del espirograma, multiplicando el FEV1 (en lt) x 35 o 40, según los autores.
Esta fórmula sirve tanto para los individuos sanos como para los enfermos con
trastornos respiratorios obstructivos y restrictivos.
Si se observa una MVV muy
baja en un usuario que parece cooperar de forma activa, hay que pensar en una
debilidad neuromuscular. Exceptuando los casos de enfermedad neuromuscular muy
avanzada, la mayoría de los usuarios son capaces de efectuar un esfuerzo
respiratorio aislado como un FVC. La MVV requiere un esfuerzo mucho mayor, y su
alteración demuestra la existencia de músculos respiratorios débiles y
fatigables. La MVV disminuye progresivamente cuando existe un aumento de la
debilidad de los músculos respiratorios; junto con las presiones inspiratoria y
espiratoria máximas, la MVV es en ocasiones, la única prueba funcional
respiratoria anómala en ciertos individuos con una enfermedad neuromuscular
relativamente grave.
La MVV es importante
también en la valoración del riesgo quirúrgico, pues refleja la gravedad de la
obstrucción de las vías aéreas y también las reservas respiratorias, la fuerza
muscular y el grado de motivación del usuario. Volúmenes pulmonares, valor de
volúmenes, capacidad total, capacidad residual funcional, capacidad reserva
respiratoria.
3
COLAPSO PULMONAR
El colapso pulmonar ocurre
cuando el aire escapa del pulmón. El aire luego llena el espacio por fuera del
pulmón, entre éste y la pared torácica. Esta acumulación de aire ejerce presión
sobre el pulmón, así que éste no se puede expandir tanto como lo hace
normalmente cuando usted inspira.
El término médico para esta
afección es neumotórax.
Causas
El colapso pulmonar puede
ser causado por una lesión al pulmón. Las lesiones pueden abarcar herida por
arma de fuego o cuchillo en el tórax, fractura de una costilla o ciertos
procedimientos médicos.
En algunos casos, un
colapso pulmonar es causado por ampollas de aire (vesículas) que se rompen,
enviando aire hacia el espacio que rodea el pulmón. Esto puede resultar de los
cambios de presión del aire como sucede al bucear o viajar a grandes alturas.
Las personas altas y
delgadas y los fumadores tienen mayor probabilidad de sufrir un colapso
pulmonar.
Las neumopatías también
pueden incrementar la posibilidad de sufrir un colapso pulmonar y son, entre
otras:
·
Asma
·
EPOC
·
Fibrosis
quística
·
Tuberculosis
·
Tos
ferina
En algunos casos, un
colapso pulmonar ocurre sin ninguna causa. Esto se denomina neumotórax
espontáneo o atelectasia pulmonar.
La ventilación mecánica es
una estrategia terapéutica que consiste en remplazar o asistir mecánicamente la
ventilación pulmonar espontánea cuando ésta es inexistente o ineficaz para la
vida. Para llevar a cabo la ventilación mecánica se puede recurrir o bien a una
máquina (ventilador mecánico) o bien a una persona bombeando el aire
manualmente mediante la compresión de una bolsa o fuelle de aire.
Se llama ventilación
pulmonar al intercambio de gases entre los pulmones y la atmósfera. Tiene como
fin permitir la oxigenación de la sangre (captación de oxígeno) y la
eliminación de dióxido de carbono.
En la ventilación
espontánea, durante la inspiración, un individuo genera presiones
intratorácicas negativas al aumentar el volumen torácico gracias a la
musculatura respiratoria (principalmente el diafragma). La presión en el
interior del tórax se hace menor que la atmosférica, generando así un gradiente
de presiones que provoca la entrada de aire a los pulmones para equilibrar esa
diferencia. La espiración (salida de aire) normalmente es un proceso pasivo.
Durante la ventilación
espontánea se introduce y expulsa un volumen regular de aire llamado volumen
tidal, de aproximadamente ½ litro, a una frecuencia respiratoria determinada
(12 – 20 respiraciones por minuto).
3.1
MECANISMOS
QUE CAUSAN UN COLAPSO PULMONAR
Son dos
mecanismos
•
La presión
intrapleural negativa ayuda a mantener los pulmones distendidos.
•
La sustancia tensioactiva o surfactante disminuye la
tensión superficial de los liquídos que rodean a los alvéolos. La sustancia
tensioactiva es una mezcla bifásica de proteínas fosfolípidos e iones ;
«dipalmitoil- lecitina» (apoproteiina surfactante e iones de calcio). Previene
el edema pulmonar, es producido por el epitelio alveolar las células tipo II.
3.2
MECANISMOS QUE LLEVAN A UN
COLAPSO PULMONAR
Depende de dos
factores
•
Las fibras elásticas del
parénquima pulmonar.
•
La tensión superficial de
los líquidos que recubren a los alvéolos.
1
VOLÚMENES Y CAPACIDADES
PULMONARES
En
primer lugar se diferencian 4 niveles:
b) Nivel
de final de inspiración normal.
c) Nivel
de inspiración máxima.
d) Nivel
de espiración máxima.
Convencionalmente
las cantidades de aire comprendidas entre dos niveles contiguos se denominan
volúmenes, y la suma de dos o más de estos, capacidades. Se distinguen 4
volúmenes:
1.
Volumen corriente (VC): cantidad de
aire que entra en una inspiración o sale en una espiración, en las condiciones
de actividad que se especifiquen (reposo, ejercicio).
2.
Volumen de reserva inspiratoria (VRI):
cantidad máxima de aire que se puede inspirar por sobre el nivel de inspiración
espontánea de reposo.
3.
Volumen de reserva espiratoria (VRE):
máxima cantidad de aire que se puede expulsar a partir del nivel espiratorio
espontáneo normal.
4.
Volumen residual (VR): cantidad de
aire que queda en el pulmón después de una espiración forzada máxima. Este
volumen no puede medirse con el espirómetro.
Las
capacidades son también 4:
1.
Capacidad pulmonar total (CPT):
cantidad de gas contenido en el pulmón en inspiración máxima. Corresponde a la
suma de los cuatro volúmenes ya descritos.
2. Capacidad vital (CV): cantidad total
de aire movilizado entre una inspiración y espiración máximas. Incluye el
volumen corriente y los volúmenes de reserva inspiratoria y espiratoria.
3.
Capacidad inspiratoria (CI): máximo
volumen de gas que puede inspirarse a partir de una espiración normal.
Comprende los volúmenes corriente y de reserva inspiratoria.
4.
Capacidad residual funcional (CRF):
volumen de gas que permanece en el pulmón al término de la espiración normal;
representa la suma del volumen residual y volumen de reserva espiratoria.
Estas
subdivisiones tienen una significación fisiológica que pasaremos a analizar en
sus principales aspectos.
1.1
VOLUMEN CORRIENTE
En
los adultos, en la respiración espontánea en reposo se inspiran y espiran en
cada ciclo respiratorio entre 400 y 600 ml, cantidad que se repite en forma
bastante regular y se denomina volumen corriente, por ser el que se mueve o
corre. Esta cantidad es aproximadamente sólo una décima parte de lo que el
pulmón puede movilizar, existiendo, por lo tanto, importantes reservas de
inspiración y espiración, a las cuales se recurre cuando aumentan las demandas
por ejercicio físico, fonación, risa, llanto, etc.
1.2
CAPACIDAD VITAL
Esta
capacidad está constituida por la suma del volumen corriente y las reservas
inspiratoria y espiratoria. Representa el máximo de aire que se puede movilizar
en una sola maniobra respiratoria. En 1846, John Hutchinson desarrolló el
método de medición aún vigente y sentó las bases para su aplicación clínica.
Por estimar que revelaba la potencialidad de vida del individuo la denominó
capacidad vital.
La
capacidad vital se mide directamente en un espirómetro, y los valores
encontrados se expresan directamente en litros o mililitros y como porcentaje
de un valor teórico predeterminado o de referencia, que depende de la talla,
edad y sexo del individuo.
Es
importante tener presente que el valor de referencia es un promedio con
márgenes de más/menos 20 a 25%, lo que puede conducir a serios errores de
interpretación. Supongamos el siguiente ejemplo: una persona normal que, de
haber sido examinado cuando estaba sano, hubiera tenido una CV de 120% del
valor teórico promedio, presenta una enfermedad pulmonar que reduce su CV a un
85% del mismo promedio teórico. Este último valor deberá ser informado
como "dentro de los límites normales", aunque para el paciente
significa una pérdida de un tercio de su capacidad vital. Por esta razón es
importante instruir a la persona sana que se hace una espirometría en un
examen de salud o en un examen pre-ocupacional, que guarde siempre sus
resultados, ya que así contará con un valor de referencia personal.
La
CV depende de la correcta integración entre la generación y la conducción de
los estímulos respiratorios, de la capacidad muscular respiratoria, de la
mecánica esquelética y del estado del pulmón. El nivel de inspiración máxima,
límite superior de la CV, no está determinado por impedimentos mecánicos sino
por reflejos propioceptivos generados en el pulmón distendido, que frenan la
contracción muscular. Esto explica que en el cadáver con el tórax abierto, éste
pueda distenderse hasta un mayor volumen.
Dada
la amplia reserva del fuelle, las alteraciones funcionales leves suelen pasar
inadvertidas para el paciente, pero pueden ser captadas en la medición de la
CV. Esta puede disminuir por múltiples mecanismos, que pueden reducirse a dos
tipos fundamentales: los trastornos obstructivos que reducen la CV por aumento
del volumen residual atrapado en el pulmón y los trastornos restrictivos que,
como su nombre lo indica, restringen el volumen del pulmón utilizable, debido a
ocupación o colapso de alvéolos, infiltración del intersticio, ocupación del
espacio pleural, restricciones a la movilidad del tórax, debilidad muscular,
etc. Al referirnos a la fisiopatología del aparato respiratorio analizaremos
estos aspectos con mayor detalle.
1.3
VOLUMEN RESIDUAL Y CAPACIDAD
RESIDUAL FUNCIONAL
El
volumen residual (VR) es el aire que queda en el pulmón después de una
espiración forzada máxima, por lo que no se puede medir en la espirometría,
debiendo recurrirse a métodos indirectos de mayor complejidad. Sumado al
volumen de reserva espiratoria, constituye la capacidad residual funcional
(CRF), que es la cantidad de gas que se mantiene en el pulmón al final de
la espiración espontánea. Cumple diversas funciones:
a)
Permite que la composición del aire alveolar oscile muy levemente, ya que los 2
a 3 litros de gas que permanecen en el pulmón diluyen el aire inspirado,
impidiendo cambios bruscos en la composición del aire alveolar. Si el aire
alveolar se recambiara totalmente por aire atmosférico, el CO2 de la
sangre venosa al llegar al alvéolo se liberaría explosivamente en forma de
burbujas y se producirían cambios bruscos y violentos en el equilibrio ácido
base.
b)
Sirve como reservorio de oxígeno, lo que permite que la sangre siga removiendo
este gas del pulmón en forma continua durante la espiración y en períodos
cortos de apnea.
c)
Mantiene un volumen de aire en los alveolos que impide su colapso, situación
que exigiría generar grandes presiones para volver a expandirlos La
capacidad residual funcional está determinada por la interacción de las fuerzas
elásticas del pulmón, que tienden al colapso, y las del tórax, que tienden a la
expansión. Su posición de equilibrio corresponde al nivel de final de
espiración en reposo.
Para
llegar al volumen residual la espiración forzada tiene que vencer la
elasticidad torácica, siendo finalmente limitada por reflejos propioceptivos
toracopulmonares y por el cierre de las pequeñas vías aéreas. Este último
fenómeno se debe a que la disminución del volumen pulmonar reduce la tracción
elástica que el parénquima pulmonar ejerce sobre los bronquíolos,
manteniéndolos abiertos. Por el envejecimiento normal de los elementos
elásticos del pulmón, este fenómeno de cierre se acentúa con la edad, con lo
que el VR aumenta, representando una fracción progresivamente mayor de la
capacidad pulmonar total (30% hasta los 35 años y 40% sobre los 50 años).
En
cifras absolutas, el VR de un hombre de 20 años, 1,70 m de estatura, con una
CPT de 6 L, es de aproximadamente 1,8 L.
1.4
CAPACIDAD INSPIRATORIA
Cuando
el volumen residual y la capacidad residual funcional, de compleja
medición,aumentan en forma aguda sin modificar la capacidad pulmonar total, lo
hacen a expensas de la capacidad inspiratoria, medible en la esprometria. Esto
permite la evaluacion indirecta fácil de la capacidad residual funcional en el
fenómeno de hiperinflacion dinámica, de gran importancia en en la
obstruccion bronquial difusa

Figura: Volúmenes y
capacidades pulmonares. Los niveles de inspiración máxima, reposo inspiratorio,
reposo espiratorio, espiración máxima determinan los volúmenes de reserva
inspiratoria (VRI), corriente (VC), de reserva espiratoria (VRE) y residual
(VR). La suma de distintos volúmenes resulta en las capacidades inspiratorias
(CI), residual funcional (CRF), vital (CV) y pulmonar total (CPT).
2 IMPORTANCIA DEL VOLUMEN RESIDUAL
La conservación de un cierto
volumen de aire en las vías respiratorias cuando ya no somos capaces de
expulsar más aire en la espiración forzada, es esencial para mantener un
equilibrio en la presión interna de los alvéolos, aspecto vital para que los
pulmones puedan mantener su actividad con normalidad.
Si nuestros pulmones no
conservaran permanentemente un cierto volumen de aire residual, los alvéolos se
vaciarían normalmente, acabando aplastados y con ello colapsados por el aumento
de la presión de succión que se produce en su interior para compensar este
vacío.
Este volumen de aire residual
oscila entre 1 y 1,2 litros de aire según las personas.
A modo de conclusión, los
volúmenes respiratorios nos dan información sobre la cantidad de aire que el
individuo es capaz de movilizar en relación con los distintos tipos de esfuerzo
respiratorio, es decir el aire que se moviliza sin esfuerzo, el que se inhala
de manera aislada al forzar la inspiración, o el que se puede expulsar
exclusivamente en la espiración forzada.
De este modo, gracias a los
volúmenes respiratorios podemos saber si nuestros mecanismos respiratorios
fisiológicos funcionan adecuadamente y rinden de acuerdo a sus posibilidades.
El rendimiento de algunos de
estos volúmenes puede ser aumentado a través de un cierto entrenamiento
respiratorio. Así la relajación de los músculos torácicos y la flexibilización
de la caja torácica nos pueden ayudar a aumentar la amplitud de la inspiración,
es decir el volumen de reserva inspiratoria (VRI).
Así mismo, el entrenamiento de
los músculos espiratorios nos puede ayudar a expulsar una mayor cantidad de
aire, mejorando de este modo el volumen de reserva espiratorio (VRE).
El volumen residual (VR) es el aire que
queda en el pulmón después de una espiración forzada máxima, por lo que no se
puede medir en la espirometría, debiendo recurrirse a métodos indirectos de
mayor complejidad. Sumado al volumen de reserva espiratoria, constituye la
capacidad residual funcional (CRF), que es la cantidad de gas que se mantiene
en el pulmón al final de la
espiración espontánea. Cumple
diversas funciones:a) Permite que la composición del aire alveolar oscile muy levemente, ya que los 2 a 3 litros de gas que permanecen en el pulmón diluyen el aire inspirado, impidiendo cambios bruscos en la composición del aire alveolar. Si el aire alveolar se recambiara totalmente por aire atmosférico, el CO2 de la sangre venosa al llegar al alvéolo se liberaría explosivamente en forma de burbujas y se producirían cambios bruscos y violentos en el equilibrio ácido base.
b) Sirve como reservorio de oxígeno, lo que permite que la sangre siga removiendo este gas del pulmón en forma continua durante la espiración y en períodos cortos de apnea.
c) Mantiene un volumen de aire en los alveolos que impide su colapso, situación que exigiría generar grandes presiones para volver a expandirlos La capacidad residual funcional está determinada por la interacción de las fuerzas elásticas del pulmón, que tienden al colapso, y las del tórax, que tienden a la expansión. Su posición de equilibrio corresponde al nivel de final de espiración en reposo.
Para llegar al volumen residual la espiración forzada tiene que vencer la elasticidad torácica, siendo finalmente limitada por reflejos propioceptivos toracopulmonares y por el cierre de las pequeñas vías aéreas. Este último fenómeno se debe a que la disminución del volumen pulmonar reduce la tracción elástica que el parénquima pulmonar ejerce sobre los bronquíolos, manteniéndolos abiertos. Por el envejecimiento normal de los elementos elásticos del pulmón, este fenómeno de cierre se acentúa con la edad, con lo que el VR aumenta, representando una fracción progresivamente mayor de la capacidad pulmonar total (30% hasta los 35 años y 40% sobre los 50 años).
En cifras absolutas, el VR de un hombre de 20 años, 1,70 m de estatura, con una CPT de 6 L, es de aproximadamente 1,8 L.
3
FORMAS QUÍMICAS EN QUE SE
TRANSPORTA EL CO2
El CO2 se produce
a nivel las mitocondrias, como producto final del metabolismo celular. Desde
las mitocondrias atraviesa el citoplasma, pasa a la sangre en los capilares
tisulares y es llevado por la sangre al alvéolo, desde donde se elimina a la
atmósfera gracias a la ventilación alveolar.
El CO2 es
transportado en la sangre, tanto en combinaciones químicas como en solución
física. Debido a que tiene la propiedad de hidratarse, formando ácido carbónico
en una reacción reversible, el transporte de CO2 y su
eliminación son parte fundamental del equilibrio ácido-base. La cantidad total
de CO2 en la sangre arterial es de aproximadamente 48 volúmenes
en 100ml de sangre.
Transporte
en el plasma:
Se realiza en tres formas:
1.- Parte se mantiene
disuelta físicamente en el plasma, dependiendo de la presión parcial de CO2 y
de su coeficiente de solubilidad.
2.- Otra parte forma
compuestos carbamínicos con las proteínas plasmáticas en una reacción rápida
que no requiere de catalizador:
R-NH2 +
CO2  R-NHCOO- +
H+
R-NHCOO- +
H+
3.- Una pequeña cantidad
reacciona con el agua para formar ácido carbónico e implicarse en el equilibrio
ácido-base:
CO2 +
H2O H2CO3 H+ + HCO3-
Transporte
por el glóbulo rojo
La mayor parte del CO2 que
difunde desde los tejidos hacia los capilares entra al glóbulo rojo, donde se
transporta en las siguientes formas:
1.-Una pequeña fracción
permanece disuelta en el líquido dentro del glóbulo
2.-Parte del CO2 se
combina con los grupos amino de la hemoglobina para formar compuestos
carbamínicos.
3.-La mayor parte del CO2 que
penetra al glóbulo rojo a nivel tisular se hidrata como en el plasma, pero a
mayor velocidad, ya que en el eritrocito existe una alta concentración de la
enzima anhidrasa carbónica que cataliza la reacción. El bicarbonato que se
forma se disocia en H+y HCO3. Los iones H+son
captados por la hemoglobina y los aniones HCO3 salen del
glóbulo rojo hacia el plasma, donde la concentración de este ión es menor,
intercambiándose por el anión cloro (efecto Hamburger).
Al entregar O2 la Hb oxigenada se transforma en Hb reducida, que por ser un ácido débil tiene poder tamponador de H+. Con ello aumenta la capacidad de transporte de CO2 (efecto Haldane). De este modo, la entrega de O2 y la captación de CO2 que tienen lugar en los capilares sistémicos son dos procesos que se favorecen mutuamente: un aumento de la presión de CO2 en la sangre capilar, con la consiguiente disminución del pH, que facilita la entrega de O2 (efecto Bohr), a la par que el aumento de Hb reducida facilita la captación de CO2 (efecto Haldane).
Al entregar O2 la Hb oxigenada se transforma en Hb reducida, que por ser un ácido débil tiene poder tamponador de H+. Con ello aumenta la capacidad de transporte de CO2 (efecto Haldane). De este modo, la entrega de O2 y la captación de CO2 que tienen lugar en los capilares sistémicos son dos procesos que se favorecen mutuamente: un aumento de la presión de CO2 en la sangre capilar, con la consiguiente disminución del pH, que facilita la entrega de O2 (efecto Bohr), a la par que el aumento de Hb reducida facilita la captación de CO2 (efecto Haldane).
En los pulmones, la PCO2 alveolar (40 mmHg) es menor que la PCO2 de la sangre venosa (46 mmHg) y las reacciones antes descritas se desplazan hacia la izquierda debido a que parte del CO2 difunde desde la sangre al alvéolo. Al mismo tiempo, la Hb reducida se oxigena, transformándose en un ácido más fuerte, lo que significa liberar CO2 del glóbulo rojo. Simultáneamente, el bicarbonato del plasma entra al eritrocito donde forma H2CO3, que se disocia en CO2 y H2O en presencia de la enzima anhidrasa carbónica. El CO2 formado difunde a través de la membrana del eritrocito al plasma, atraviesa la membrana alvéolo-capilar y es eliminado con la ventilación.

1 UNIDAD RESPIRATORIA
La unidad respiratoria está compuesta por
bronquiolos respiratorios, conductos alveolares, los atrios y los alvéolos. Las
paredes alveolares son extremadamente delgadas, y en su interior existe una red
casi sólida de capilares interconectados. Por tanto, es obvio que los gases
alveolares están muy próximos a la sangre de los capilares; el intercambio
gaseoso entre el aire alveolar y la sangre pulmonar, se produce a través de las
membranas de todas las porciones terminales de los pulmones, no meramente en
los propios alvéolos. Estas membranas se denominan colectivamente membrana respiratoria,
llamada también membrana pulmonar.
1.1
MEMBRANA RESPIRATORIA
Está
constituida por las siguientes capas
·
Una capa de líquido que reviste el alvéolo y
que contiene el agente tensión-activo que disminuye la tensión superficial del
líquido alveolar:
·
El epitelio alveolar compuesto de células
epiteliales finas.
·
Una membrana basal epitelial.
·
Un espacio intersticial fino entre el
epitelio alveolar y la membrana capilar.
·
Una membrana basal del capilar que en muchos
lugares se fusiona con la membrana basal epitelial.
·
La membrana endotelial capilar.
A pesar de
la cantidad de capas presentes, la membrana respiratoria tiene un espesor de
tan solo 0,2 micras en algunos lugares, y es en promedio de 0,6 micras.
El
diámetro del capilar es de tan solo micras, por lo que el hematíe debe
deformarse, y su membrana se pone en contacto con la pared del capilar,
entonces del Co2 no precisa atravesar cantidades significativas de plasma
cuando difunde. Esto aumenta la velocidad de la difusión.
Factores
que afectan la tasa de difusión de gases a través de la membrana respiratoria
·
El espesor
de la membrana respiratoria: Debido a que la tasa de difusión es
inversamente proporcional al espesor de la membrana, cualquier factor que
aumenta el espesor a más de dos o tres veces el normal puede interferir
significativamente con el intercambio de gases de la respiración normal;
·
El área de
la superficie de la membrana respiratoria: Cuando la superficie
total está disminuida entre un tercio y un cuarto de lo normal, el intercambio
de gases a través de la membrana está significativamente interferido, incluso
en condiciones de reposo;
·
El
coeficiente de difusión de la transferencia de cada gas (solubilidad): Para una
diferencia de presión dada, el dióxido de carbono difunde a través de la membrana
20 veces más rápidamente que el oxígeno. El oxígeno a su vez difunde dos veces
más rápidamente que el nitrógeno. La diferencia de presión a través de la
membrana respiratoria es la diferencia entre la presión parcial del gas en los
alvéolos y la presión del gas en la sangre.
Captación
de oxígeno por la sangre
La Po2
gaseoso del alvéolo es de 104 mm de Hg en promedio, mientras que la Po2 de la
sangre venosa que penetra en el capilar sólo es de 40 mm de Hg, debido a que
una gran cantidad de oxígeno ha sido extraída de esta sangre a su paso por los
tejidos periféricos. Por tanto, la diferencia de presión inicial que hace que
el oxígeno difunda al capilar pulmonar es de 104 - 40, ó 64 mm de Hg. O sea, la
Po2 se eleva hasta un nivel prácticamente igual al del aire alveolar cuando la
sangre ya ha recorrido un tercio del trayecto capilar, alcanzando casi 104 mm
de Hg.
Transporte de oxígeno en la sangre
Aproximadamente
el 98% de la sangre que penetra en la aurícula izquierda procedente de los
pulmones ha pasado por los capilares alveolares y se ha oxigenado hasta una Po2
de unos 104 mm de Hg. Otro 2% de la sangre ha pasado directamente desde la
aorta a través de la circulación bronquial, que riega fundamentalmente los
tejidos profundos de los pulmones y no está expuesta al aire pulmonar. Este
flujo de sangre representa al flujo de cortocircuito, lo que significa que
la sangre ha pasado de largo por las zonas de intercambio gaseoso. Esta sangre
de cortocircuito tiene una Po2 de unos 40 mm de Hg igual a la de la sangre
venosa. Esta sangre se combina en las venas pulmonares con la sangre oxigenada
procedente de los capilares pulmonares; esta mezcla de las sangres se denomina
adición de sangre venosa, y hace que la Po2 de la sangre bombeada por la mitad
izquierda del corazón hacia la aorta descienda a unos 95 mm de Hg.
Difusión de oxígeno de los capilares
periféricos al líquido tisular
Cuando la
sangre arterial alcanza los tejidos periféricos, su Po2 en los capilares siguen
siendo 95 mm de Hg. Por otra parte, la Po2 del líquido intersticial que rodea
las células de los tejidos sólo es de unos 40 mm de Hg. Por tanto existe una
diferencia de presión que hace que el oxígeno difunda rápidamente de la sangre
a los tejidos, haciendo que la Po2 capilar descienda hasta igualar casi los 40
mm de Hg de presión del intersticio. Por tanto, la Po2 de la sangre que
abandona los capilares tisulares y penetra en las venas es también de unos 40
mm de Hg.
Difusión de oxígeno de los capilares a las
células
Las
células siempre están utilizando oxígeno. Por lo tanto, la Po2 intracelular
siempre es menor que la Po2 capilar. Además, en muchos casos existe una
distancia considerable entre los capilares y las células. Por tanto, la Po2
intracelular normal varía entre cifras tan bajas como 5 mm de Hg y tan elevadas
como 40 mm de Hg, siendo el promedio de 23 mm de Hg. Debido a que normalmente
sólo se requiere de 1 a 3 mm de Hg de presión de oxígeno para el soporte de los
procesos químicos que consumen oxígeno en la célula, se puede ver que incluso esta
baja Po2 celular de 23 mm de Hg es más que adecuada y brinda un gran factor de
seguridad.
Difusión del dióxido de carbono desde las
células de los tejidos a los capilares tisulares, y de los capilares pulmonares
a los alvéolos
Cuando las
células utilizan oxígeno, en su mayor parte se convierte en dióxido de carbono,
y esto aumenta la Pco2 intracelular. Por tanto, el dióxido de carbono difunde
de las células a los capilares tisulares y después es transportado, por la
sangre a los pulmones. En los pulmones, difunde desde los capilares pulmonares
a los alvéolos. Existe una importante diferencia entre la difusión de dióxido
de carbono y la del oxígeno: el dióxido de carbono puede difundir unas 20 veces
más deprisa que el oxígeno. Por tanto, las diferencias de presión que producen
la difusión de dióxido de carbono son, en todos los casos, mucho menores que
las diferencias de presión necesarias para producir la difusión del oxígeno.
Estas presiones son:
·
Pco2 intracelular de unos 46 mm de Hg; Pco2
intersticial de unos 45 mm de Hg; por tanto, sólo existe una diferencia de
presión de 1 mm de Hg;
·
Pco2 de la sangre arterial que penetra en los
tejidos de 40 mm de Hg; Pco2 de la sangre venosa que abandona los tejidos, de
unos 45 mm de Hg; por tanto, la sangre capilar tisular entra en equilibrio casi
exacto con la Pco2 intersticial, que también es de 45 mm de Hg;
·
Pco2 de la sangre venosa que entra en los
capilares pulmonares es de 45 mm de Hg; pco2 del aire alveolar, 40 mm de Hg;
por tanto, sólo una diferencia de presión de 5 mm de Hg produce toda la
difusión de dióxido de carbono necesaria, desde los capilares pulmonares a los
alvéolos. Además, la Pco2 de la sangre capilar pulmonar disminuye hasta igualar
casi exactamente la Pco2 alveolar de 40 mm de Hg antes de haber recorrido más
de un tercio de la longitud de los capilares.
1.2
REGULACIÓN DE LA RESPIRACIÓN

1.1
CENTRO RESPIRATORIO
El centro
respiratorio está compuesto de varios grupos de neuronas localizadas
bilateralmente en el bulbo raquídeo y en la protuberancia. Está dividido en
tres grupos principales de neuronas:
·
Grupo respiratorio dorsal, localizado en la
porción dorsal del bulbo, que origina principalmente la inspiración
·
Grupo respiratorio ventral, localizado en la
parte ventrolateral del bulbo, que puede originar la espiración o la
inspiración, dependiendo de qué neuronas del grupo se estimulen
·
Centro neumotáxico, localizado dorsalmente en
la parte superior de la protuberancia, que ayuda a controlar la frecuencia y el
patrón respiratorio.
Grupo respiratorio dorsal de neuronas: sus
funciones inspiratorias y rítmicas
Todas o la
mayoría de sus neuronas están localizadas dentro del núcleo del fascículo
solitario. El núcleo del fascículo solitario es también la terminación
sensitiva de los nervios vago y glosofaríngeo, que transmiten al centro
respiratorio señales sensitivas de los quimiorreceptores periféricos, los
barorreceptores, y varios tipos de receptores del interior del pulmón.
Señal de rampa inspiratoria
La señal
nerviosa que se transmite a los músculos inspiratorios primarios como el
diafragma no es una salva instantánea de potenciales de acción. Por el
contrario, en la respiración normal, la inspiración comienza débilmente y crece
en forma de rampa durante un período de 2 segundos. Cesa repentinamente durante
los 3 segundos siguientes, con lo que cesa la estimulación del diafragma y
permite que la retracción elástica de la pared torácica y los pulmones origine
la espiración. Por tanto, la señal inspiratoria es una señal en rampa. La
ventaja evidente de esto es que produce un aumento sostenido del volumen de los
pulmones durante la inspiración, en vez de boqueadas inspiratorias.
Existen
dos formas de control de la rampa inspiratoria:
·
Control de la tasa de incremento de la señal
de la rampa, de forma que durante la respiración activa la rampa aumenta
rápidamente y por tanto llena los pulmones también con rapidez.
·
Control del punto límite en el cual cesa
repentinamente la rampa. Esta es la forma habitual de controlar la frecuencia
respiratoria: es decir, cuanto antes cesa la rampa, menos dura la inspiración.
2
REGULACIÓN DE LA ACTIVIDAD DEL
CENTRO RESPIRATORIO
Un incremento en la PCO2,
en la concentración de h+ de la sangre arterial o una caída de PO2,
aumentan la actividad del centro respiratorio, y los cambios en la dirección
opuesta tienen un efecto inhibidor ligero. Los efectos de las variaciones en la
química sanguínea sobre la ventilación están mediados por los quimiorreceptores respiratorios,
células receptoras en el bulbo y en los cuerpos carotídeos y aórticos,
sensibles a los cambios en la composición de la sangre, que inician impulsos
que estimulan el centro respiratorio. Además del control
químico respiratorio básico, otros aferentes
proveen controles no químicos para los "ajustes finos" que afectan la
respiración en situaciones particulares.
Generan el ritmo
respiratorio basal, procesan la información de los sensores y modifican, en
consecuencia, su nivel de actividad.
1. Los controladores o
centros respiratorios tienen las siguientes funciones:
2. Establecer el ritmo
de la respiración y actuar como generadores centrales del patrón
respiratorio.
3. Transmitir ese ritmo
central a las motoneuronas que inervan los músculos respiratorios.
4. Ajustar el ritmo
respiratorio y de la respuesta motora a las necesidades metabólicas (funciones
homeostáticas), así como para cubrir las funciones conductuales y voluntarias
(funciones no homeostáticas).
5. Utilizar el mismo
gasto de energía para llevar a cabo varias funciones.
Los experimentos de
transección a distintos niveles del SNC permitieron concluir que los centros
encargados del control automático del ritmo respiratorio se localizaban en el
tronco encefálico; en función estos resultados se hablaba de:
·
Centro neumotáxico, parte rostral de la protuberancia
·
Centro apnéustico, en la parte ventral
·
Serie de centros bulbares (principales responsables del ritmo
respiratorio)
Los centros
neumotáxico y apnéustico (o centros suprabulbares) se encargan de modular y
afinar el centro respiratorio.
·
Los centros bulbares
Los estudios electrofisiológicos
han mostrado la existencia de varios grupos neuronales en distintos núcleos
bulbares, capaces de aumentar su actividad (frecuencia de disparo de
potenciales de acción) durante la inspiración; sin embargo, a diferencia de lo
que ocurre en el corazón, no parece que haya un grupo único de células
marcapasos en el bulbo donde se origina el ritmo respiratorio básico; por el
contrario, el patrón de inspiración-espiración es generado neuronas
interconectadas, las cuales forman redes que actúan como circuitos oscilantes.
Durante la
inspiración, entre dichas redes, la frecuencia de disparo aumenta en varias
células (en distintos puntos), mientras que en la espiración otros grupos se
activan.
Las neuronas que
constituyen el CPG, se localizan de forma más o menos difusa bilateralmente en
el bulbo y forman parte de, al menos, 2 grupos de núcleos: respiratorio dorsal
y respiratorio ventral.
Grupo respiratorio dorsal: Está formado por neuronas localizadas
en la región dorso medial del bulbo y forma parte del núcleo del tracto
solitario (nTS). Contiene fundamentalmente neuronas inspiratorias de distintos
tipos, clasificadas teniendo en cuenta el momento de la inspiración en el que
aumenta su actividad y el patrón de esta.
Funciones: Envían proyecciones a las motoneuronas de los nervios frénicos e
intercostales y son, por tanto, las responsables de la actividad mantenida del
diafragma durante la inspiración; también establecen conexiones con el grupo
respiratorio ventral.
Núcleo del tracto solitario
Constituye la
principal proyección de vías aferentes viscerales de los nervios glosofaríngeo
y vago, que llevan informaciones de la PO2, PCO2 y
el pH (proveniente de los quimiorreceptores periféricos) y de la presión
arterial sistémica (desde los barorreceptores aórticos).
El vago traslada
información desde los receptores de estiramiento pulmonar, de modo que la
localización del grupo respiratorio dorsal en el núcleo del tracto solitario,
indica que es el lugar de integración de muchos reflejos cardiopulmonares que
afectan el ritmo respiratorio.
Grupo respiratorio ventral (GRV): Su distribución anatómica es más
difusa que la del dorsal y está constituido por agregados de células que se
extienden longitudinalmente por el bulbo, desde su zona caudal hasta la más rostral.
Se puede dividir en
tres regiones:
- Parte caudal,
denominada núcleo retroambiguo (GRV caudal o nRA), por su relación con el
núcleo ambiguo (nA) contiene fundamentalmente neuronas espiratorias. Las zonas
de muchas de estas neuronas establecen sinapsis con las motoneuronas que
controlan los músculos espiratorios intercostales y abdominales (espiración
forzada).
- Parte intermedia,
denominada núcleo paraambiguo (GRV intermedio o nPA). Por su distribución
paralela al núcleo ambiguo contiene fundamentalmente neuronas inspiratorias,
pero incluye también las propiobulbares, las cuales coordinan la actividad de
los músculos respiratorios con el control de la resistencia de las vías aéreas
superiores y desempeñan una función clave dentro del CPG.
- Parte más
rostral (GVR rostral), se localiza en la vecindad del núcleo
retrofacial (nRF) e incluye una densa población de neuronas que se agrupan
y forman el llamado complejo de Bötzinger.
-
Constitución del complejo de Bötzinger
Está formado por
diversos tipos funcionales de neuronas espiratorias, algunas motoneuronas que
inervan la laringe y la faringe, otras son interneuronas.
Recientemente ha sido
identificado el complejo de pre_Bötzinger, pues en esta zona se localiza el
CPG, ya que es capaz de generar un ritmo respiratorio, incluso en preparaciones
aisladas, y su lesión da lugar a alteraciones del ritmo, tanto in vivo como in vitro.
-
Complejo de pre-Bötzinger
Contiene hasta 6
tipos de neuronas respiratorias, que debido a sus propiedades intrínsecas y a
las interacciones sinápticas que establecen, permiten generar y mantener una
actividad cíclica espontánea en forma de salvas de disparos de potenciales de
acción; observaciones que indican su función esencial en la génesis del ritmo
respiratorio.
·
El centro apnéustico
Su localización
hística aún no está bien precisada, pero parece estar formado por una red
neuronal difusa, ubicada en la formación reticular de la protuberancia.
En investigaciones
más recientes se precisa que la ablación del centro neumotáxico, al combinarse
con la vagotonía, da lugar a una respiración con inspiraciones prolongadas,
separadas por espiraciones breves.
Función: Se estima que es el centro o lugar de proyección e integración de
diferentes tipos de información aferente, que pueden finalizar la inspiración
(interruptor inspiratorio); proceso identificado en inglés con las siglas IO-S
(inspiratory-off switch).
Tanto la estimulación
vagal, por el aumento del volumen pulmonar, como la del centro neumotáxico
activan las neuronas IO-S y hacen que acabe la fase de inspiración.
Cuando este mecanismo
se inactiva mediante la supresión de las aferencias vagales y de los centros
superiores aparece la apneusis.
Estas neuronas
también se estimulan por el aumento de la temperatura corporal y ocasionan la
taquipnea (aumento de la frecuencia respiratoria), mecanismo que utilizan
algunos animales para disipar calor cuando están hipertérmicos.
Al igual que el
centro neumotáxico, el IO-S no parece desempeñar una función crucial en la
génesis del ritmo respiratorio básico.
- Centro neumotáxico
Está compuesto por neuronas
que se agrupan en 2 núcleos, situados en la parte rostral de la protuberancia:
1.
Núcleo
parabraquial medial
2.
Núcleo
de Köliker-Fuse
Función: Modular los centros
respiratorios bulbares, pues la estimulación de las neuronas del neumotáxico
desactiva la inspiración, regula el volumen inspiratorio y, en consecuencia, la
frecuencia respiratoria, lo cual apunta hacia el hecho de que no parece
participar en la génesis del ritmo respiratorio, ya que puede existir un patrón
normal en su ausencia.
9.1 VITALOMETRIA
Sirve para medir ciertos
volúmenes y capacidades tales como:
·
Volúmenes
de ventilación pulmonar.
·
Volúmenes
de reserva inspiratoria.
·
Volúmenes
de reserva espiratoria.
3
SONIDO. AUDICION. ONDAS
3.1
SONIDO
El sonido (del latín sonĭtus, por analogía prosódica
con ruido, chirrido, rugido,
etcétera), en física, es cualquier fenómeno que involucre la
propagación en forma de ondas elásticas (sean audibles o no),
generalmente a través de un fluido (u otro medio elástico) que esté generando
el movimiento vibratorio de un cuerpo.
El sonido humanamente audible consiste en ondas sonoras que se producen cuando
las oscilaciones de la presión del
aire, son convertidas en ondas mecánicas en el oído humano y percibidas por el
cerebro. La propagación del sonido es similar en los fluidos, donde el sonido
toma la forma de fluctuaciones de presión.1 En los cuerpos
sólidos la propagación del sonido involucra variaciones del estado tensional
del medio.

La propagación del sonido involucra transporte de energía sin transporte de
materia,
en forma de ondas mecánicas que se propagan a través de un medio elástico sólido, líquido o gaseoso. Entre los más comunes se encuentran el aire
y el agua. No se propagan en el vacío, al contrario que las ondas
electromagnéticas. Si las vibraciones se producen en la misma dirección en la
que se propaga el sonido, se trata de una onda longitudinal y si las vibraciones
son perpendiculares a la dirección de propagación es una onda transversal.
La fonética acústica concentra su interés
especialmente en los sonidos del habla: cómo se generan, cómo se perciben, y
cómo se pueden describir gráfica o cuantitativamente.
1.1
AUDICION
La audición constituye los procesos
psico-fisiológicos que proporcionan al ser humano la capacidad de oír
1. El
sonido entra en el conducto auditivo: Las ondas sonoras se desplazan a través
del conducto auditivo y alcanzan el tímpano.
2. El
tímpano y los huesos auditivos vibran: Estas ondas sonoras hacen vibrar el
tímpano y los tres huesos (huesecillos) del oído medio.
3. El
líquido se mueve a través del oído interno: Las vibraciones se transmiten a
través del líquido del oído interno en forma de espiral y hacen moverse las
minúsculas células ciliadas de la cóclea. Las células ciliadas detectan el
movimiento y lo convierten en señales químicas para el nervio auditivo.
4. Los
nervios auditivos lo comunican al cerebro: El nervio auditivo envía la
información al cerebro mediante impulsos eléctricos, que son interpretados allí
como sonido.
1.2
ONDAS
Una onda consiste en la
propagación de una perturbación de alguna propiedad de un medio, por ejemplo, densidad, presión, campo eléctrico o campo magnético, a través de dicho
medio, implicando un transporte de energía sin transporte de
materia. El medio perturbado puede ser de naturaleza diversa como aire, agua, un trozo de metal e, incluso,
inmaterial como el vacío.
La magnitud física cuya perturbación se
propaga en el medio se expresa como una función tanto de la posición como del tiempo .
Matemáticamente se dice que dicha función es una onda si verifica la ecuación
de ondas:
.
Matemáticamente se dice que dicha función es una onda si verifica la ecuación
de ondas:
Donde  es la velocidad de propagación de la
onda. Por ejemplo, ciertas perturbaciones de la presión de un medio, llamadas sonido, verifican la ecuación anterior, aunque algunas ecuaciones no lineales también
tienen soluciones ondulatorias, por ejemplo, un solitón.
es la velocidad de propagación de la
onda. Por ejemplo, ciertas perturbaciones de la presión de un medio, llamadas sonido, verifican la ecuación anterior, aunque algunas ecuaciones no lineales también
tienen soluciones ondulatorias, por ejemplo, un solitón.
2
VELOCIDAD Y ENERGÍAS DEL SONIDO
1.1
Velocidad del sonido
La velocidad del sonido es la dinámica de
propagación de las ondas sonoras. En la atmósfera terrestre es de 343 m/s (a 20 °C de temperatura, con 50 % de
humedad y a nivel del mar). La velocidad del sonido varía en
función del medio en el que se trasmite. Dado que la velocidad del sonido varía
según el medio, se utiliza el número Mach 1 para indicarla. Así un cuerpo que se mueve en el
aire a Mach 2 avanza a dos veces la velocidad del sonido en esas condiciones,
independientemente de la presión del aire o su temperatura.
La velocidad o dinámica de propagación
de la onda sonora depende de las características del medio en el que se realiza
dicha propagación y no de las características de la onda o de la fuerza que la
genera. Su propagación en un medio puede servir para estudiar algunas
propiedades de dicho medio de transmisión.
Medios de
propagación
La velocidad del sonido varía
dependiendo del medio a través del cual viajen las ondas sonoras.
En el aire, a 0 °C, el sonido
viaja a una velocidad de 331,5 m/s (por cada grado centígrado que sube la
temperatura, la velocidad del sonido aumenta en 0,6 m/s)
1.2
Energía sonora
La energía
sonora (o energía acústica) es la energía que transmiten o
transportan las ondas sonoras. Procede de la energía vibracional del foco sonoro
y se propaga a las partículas del medio que atraviesan en
forma de energía cinética (movimiento de las
partículas), y de energía potencial (cambios de presión producidos en dicho medio opresión sonora). Al irse propagando el
sonido a través del medio, la energía se transmite a la velocidad de la onda,
pero una parte de la energía sonora se disipa en forma deenergía térmica.1 La energía acústica suele tener valores absolutos
bajos, y su unidad de medida es el julio (J). Aunque puede
calcularse a partir de otras magnitudes como la intensidad sonora, también se
pueden calcular otras magnitudes relacionadas,como la densidad o el flujo de
energía acústica.
A partir de la definición
de intensidad acústica, se puede calcular la
energía acústica que atraviesa una superficie A:
![E_{son} = \int_A \left[\int_0^{\Delta t} I_{son}(\mathbf{r},t)\ dt \right] dA](file:///C:/Users/usuario/AppData/Local/Temp/msohtmlclip1/01/clip_image002.png)
Donde:
Densidad de energía
acústica
La densidad de energía
acústica o energía acústica por unidad de volumen es la cantidad
media de energía acústica ( ε ) que corresponde a un volumen unidad. Equivale también al
cociente entre la intensidad sonora y la velocidad del sonido.2 3 La unidad de medida es el julio por metro cúbico,
J/m3
Para una onda sonora plana,
la densidad de energía se calcula a partir de la potencia cuadrática media
(PRMS), la densidad del medio (ρo) y de la velocidad.
La densidad de energía
acústica total (cinética más potencial) equivale al producto de la densidad del
medio por el cuadrado de la velocidad de las partículas en la dirección de
propagación de la onda.
2
ELEMENTOS DE UNA ONDA
·
Cresta:
La cresta es el punto de máxima elongación o máxima amplitud de onda; es decir,
el punto de la onda más separado de su posición de reposo.
·
Período ( ): El periodo es el tiempo que tarda
la onda en ir de un punto de máxima amplitud al siguiente.
): El periodo es el tiempo que tarda
la onda en ir de un punto de máxima amplitud al siguiente.
·
Amplitud ( ): La amplitud es la distancia
vertical entre una cresta y el punto medio de la onda. Nótese que pueden
existir ondas cuya amplitud sea variable, es decir, crezca o decrezca con el
paso del tiempo.
): La amplitud es la distancia
vertical entre una cresta y el punto medio de la onda. Nótese que pueden
existir ondas cuya amplitud sea variable, es decir, crezca o decrezca con el
paso del tiempo.
·
Frecuencia ( ): Número de veces que es repetida
dicha vibración por unidad de tiempo. En otras palabras, es una simple
repetición de valores por un período determinado.
): Número de veces que es repetida
dicha vibración por unidad de tiempo. En otras palabras, es una simple
repetición de valores por un período determinado.
·
Valle:
Es el punto más bajo de una onda.
·
Longitud de onda ( ): Es la distancia que hay entre el
mismo punto de dos ondulaciones consecutivas, o la distancia entre dos crestas
consecutivas.
): Es la distancia que hay entre el
mismo punto de dos ondulaciones consecutivas, o la distancia entre dos crestas
consecutivas.
·
Nodo:
es el punto donde la onda cruza la línea de equilibrio.
·
Elongación
( ): es la distancia que hay, en forma
perpendicular, entre un punto de la onda y la línea de equilibrio.
): es la distancia que hay, en forma
perpendicular, entre un punto de la onda y la línea de equilibrio.
·
Ciclo:
es una oscilación, o viaje completo de ida y vuelta.
·
Velocidad
de propagación ( ): es la velocidad a la que se propaga
el movimiento ondulatorio. Su valor es el cociente de la longitud de onda y su
período.
): es la velocidad a la que se propaga
el movimiento ondulatorio. Su valor es el cociente de la longitud de onda y su
período.
1
CUALIDADES DEL SONIDO
1.1 LA ALTURA
Es la afinación de un sonido; está determinada por
la frecuencia fundamental de las ondas sonoras (es
lo que permite distinguir entre sonidos graves, agudos o medios) medida en
ciclos por segundo o hercios (Hz).Para que los humanos podamos percibir un
sonido, éste debe estar comprendido entre el rango de audición de 20 y 20.000
Hz. Por debajo de este rango tenemos los infrasonidos y por encima los
ultrasonidos.
1.2 LA DURACIÓN
Es el tiempo durante el cual se mantiene un sonido, está
determinada por la longitud, que indica el tamaño de una onda,
que es la distancia entre el principio y el final de una onda completa (ciclo);
según esto podemos decir que por duración los sonidos puieden ser largos o
cortos. Los únicos instrumentos acústicos que pueden mantener los sonidos el
tiempo que quieran, son los de cuerda con arco, como el violín por ejemplo; los
de viento dependen de la capacidad pulmonar, y los de percusión, de los golpes.
1.3
LA INTENSIDAD
Equivale a hablar de volumen: un sonido puede ser fuerte
o débil. Es la cantidad de energía acústica que contiene un sonido. La
intensidad viene determinada por la potencia, que a su vez está determinada por
la amplitud y nos permite
distinguir si el sonido es fuerte o débil.
Los sonidos que percibimos
deben superar el umbral auditivo (0 dB) y no llegar al umbral de dolor (140
dB). Esta cualidad la medimos con el sonómetro y los resultados se expresan en
decibelios (dB) en honor al científico e inventor Alexander Graham Bell.
1.4
EL TIMBRE
Es la cualidad que permite
reconocer la fuente emisora del sonido, por ejemplo, entre la misma nota (tono)
con igual intensidad producida por dos instrumentos musicales distintos. Se
define como la calidad del sonido. cada cuerpo sonoro vibra de una forma distinta.
Las diferencias se dan no solamente por la naturaleza del cuerpo sonoro
(madera, metal, piel tensada, etc), sino también por la manera de hacerlo sonar
(golpear, frotar, rascar). Una misma nota suena distinta si la toca una flauta,
un violín, una trompeta… cada instrumento tiene un timbre que lo identifica o
lo diferencia de los demás. Con la voz sucede lo mismo. El sonido dado por un
hombre, una mujer, un/a niño/a tienen distinto timbre. El timbre nos permitirá
distinguir si la voz es áspera,dulce, ronca o aterciopelada. También influye en
la variación del timbre la calidad del material que se utilice. Así pues, el
sonido será claro, sordo, agradable o molesto.
1
LA VOZ HUMANA
La física ha establecido que para que exista sonido se requieren tres elementos: un cuerpo que vibre; un soporte físico por el
que pueda transmitirse, y una caja de resonancia que amplifique esas
vibraciones, permitiendo que sean percibidas por el oído.
La voz humana cumple con
las tres condiciones señaladas:
1. El cuerpo elástico que
vibra son las cuerdas vocales.
2. El soporte físico por el
que se transmite es el aire
3. La caja de resonancia
está formada por parte de la garganta, por la boca y por la cavidad nasal.
La voz humana está
compuesta por ondas acústicas que viajan a través del aire a la velocidad del
sonido, esto es a 1,244 Km/h (o 340 m/s). Pero esta rapidez no significa
que me pueda comunicar fácilmente con puntos distantes pues la voz humana se
atenúa rápidamente, perdiendo energía a medida que viaja. Luego de unos pocos
metros ya no podemos escuchar una conversación, por tanto es de la misma
naturaleza que el resto de ondas acústicas y esto ya se conocía desde antes de
la invención del teléfono.
La voz masculina tiene un
tono fundamental de entre 100 y 200 Hz, mientras que la voz femenina es más
aguda, típicamente está entre 150 y 300 Hz. Las voces infantiles son aún más
agudas. Sin el filtrado por resonancia que produce la cavidad buco nasal
nuestras emisiones sonoras no tendrían la claridad necesaria para ser audibles.
Ese proceso de filtrado es precisamente lo que permite generar los diversos
formantes de cada unidad segmental del habla.

1.1
RANGO DE FRECUENCIAS DE LA VOZ
HUMANA
Otra característica
importante de la voz humana es que las cuerdas vocales modulan la voz en un
amplio espectro de frecuencias que van de graves a agudos en un rango
aproximado de 20Hz a 20kHz. Todo un abanico de sonidos.

1.1
EL APARATO FONADOR HUMANO
El aparato fonador lo
componen 3 grupos de órganos diferenciados
1. Órganos de respiración (Cavidades
infraglóticas: pulmones, bronquios y tráquea).
2. Órganos de fonación (Cavidades glóticas:
laringe, cuerdas vocales y resonadores -nasal, bucal y faríngeo-).
3. Órganos de articulación
(cavidad supraglóticas: paladar, lengua, dientes, labios y glotis).
1.2
LA PRODUCCIÓN DE LA VOZ
La fonación se realiza
durante la respiración, cuando el aire contenido en
los pulmones, sale de éstos por simple
relajación de la caja torácica y el diafragma y, a través de los bronquios y la tráquea, llega a la laringe.
Hay cuatro cuerdas vocales:
- Dos superiores (bandas ventriculares), que no participan en la
articulación de la voz.
- Dos inferiores, las verdaderas cuerdas vocales,
responsables de la producción de la voz.
Las dos cuerdas
- Si se abren y se recogen a los lados, el aire pasa
libremente, sin hacer presión: respiramos.
- Si, por el contrario, se juntan, el aire choca
contra ellas, produciendo el sonido que denominamos voz.
Hay tres mecanismos básicos
de producción de voz:
- Vibración de las cuerdas que produce
los sonidos tonales o sonoros (vocales,
semivocales, nasales, etc.).
- Las interrupciones (totales o parciales) en el flujo
de aire que sale de los pulmones que da lugar a los sonidos
"sordos" (fricativas, etc.
- La combinación de vibración e interrupción, como
las oclusivas sonoras (en español 'b',
'd' y 'g').
El rango vocal lo determina la flexibilidad de las cuerdas vocales, que permite diferenciar
los distintos tipos de voces (en canto: tenor, soprano, contralto, ...), en función de la altura, intensidad y timbre.
El sonido producido en las
cuerdas vocales es muy débil; por ello, debe ser amplificado. Esta
amplificación tendrá lugar en los resonadores nasal, bucal y faríngeo, donde se
producen modificaciones que consisten en el aumento de la frecuencia de ciertos sonidos y la
desvalorización de otros.
La voz humana, una vez que
sale de los resonadores, es moldeada por los articuladores (paladar, lengua,
dientes, labios y glotis), transformándose en sonidos del habla: fonemas, sílabas, palabras. La posición concreta de los articuladores
determinará el sonido que emita la voz.
1.3
MECÁNICA VOCAL
Al inicio de la fonación,
la glotis se encuentra cerrada. A continuación se comienzan a contraer los
músculos espiratorios produciendo un aumento de presión en la zona subglótica.
Este aumento de presión va a provocar la apertura de las cuerdas vocales, permitiendo
que la corriente de aire pase a través de ellas, dejando escapar una pequeña
cantidad. Una vez ocurrido esto, las cuerdas vocales se volverán a cerrar.
De esta forma, se produce
un proceso cíclico que ocurre a la vez que la fonación. Este proceso cíclico va
a producir el sonido base, que después de modulará y articulará para dar lugar
a los diferentes sonidos gracias al efecto Bernoulli.
Efecto Bernoulli
El efecto Bernoulli es el
proceso a través del cual la corriente de aire tiende a succionar los objetos
que se encuentra a su paso. En este caso, permite que las cuerdas vocales se
aproximen. Esta proximidad se debe a la elasticidad de los músculos y ligamentos
que permite recuperar su forma, después de que sobre ella se haya ejercido una
fuerza. Los tonos medios de la voz son un sonido que se produce por la
vibración de la mucosa de la cuerda vocal. Esto es producido cuando hay una
tensión normal.
En los tonos agudos o muy
graves se produce la vibración de toda la cuerda vocal, es decir, de la mucosa
y el cuerpo de la cuerda vocal. Esta diferencia de tonos es debida a la
tensión. En los tonos agudos, las cuerdas vocales se encontrarán muy tensas,
por lo que el tejido laxo que las compone se verá disminuido, en cambio, en los
tonos graves , las cuerdas vocales están relajadas y el espacio es grande.
2 BIOFÍSICA DE LA PERCEPCIÓN AUDITIVA
Es una onda mecánica – Porque necesitan un medio
material (aire, agua, cuerpo
sólido) para su propagación. Además dicho medio debe ser elástico y no rígido
para permitir la transmisión del sonido.
Es una onda longitudinal - Porque las partículas del medio actúan en
la misma dirección en
la que se propaga la onda.
Es una onda tridimensional - Son ondas que se propagan en tres
direcciones. Las ondas tridimensionales se conocen también como ondas
esféricas, porque sus frentes de ondas son esferas concéntricas que salen de la
fuente de perturbación expandiéndose en todas direcciones.
1 LA LUZ Y EL ESPECTRO ELECTROMAGNÉTICO
La luz, como la que emite el sol o una bombilla incandescente, es una forma de energía radiante. Cuando se hace pasar a través de un prisma la luz blanca de una lámpara incandescente, la luz se separa en un espectro continuo o arcoíris de colores. Se produce el mismo fenómeno cuando la luz solar atraviesa una gota de lluvia. Los diferentes colores de la luz representan cantidades distintas de energía radiante. La luz azul, por ejemplo contiene más energía que la luz roja de la misma intensidad.
Además de la luz visible, existen otras formas de energía radiante, como los rayos gamma, la radiación ultravioleta y la radiación infrarroja. Todas estas formas de energía radiantes, o radiación electromagnética, viajan por el espacio a razón de 3.00 x 10(8) m/s: la velocidad de la luz.
Esta radiación viaja en ondas; La distancia que existe entre cada cresta de ondas consecutivas se llama longitud de onda (representada por lambda). El número de crestas que pasan por un punto determinado en 1 segundo recibe el nombre de frecuencia. La velocidad de onda se obtiene multiplicando la longitud de onda por la frecuencia. La frecuencia aumenta conforme la longitud de onda disminuye, y viceversa.
El espectro electromagnético comprende en su totalidad un extenso intervalo, que va desde el rayo gamma de alta energía, con alta frecuencia y longitudes de onda cortas, hasta las ondas de radio de baja energía, con baja frecuencia y longitudes de onda larga.

-Luz
visible
La luz visible abarca tan
solo una pequeña fracción del espectro electromagnético total. Nuestros ojos perciben
longitudes de onda que van desde aproximadamente 400 nm (Luz violeta) hasta
alrededor de 750 nm (luz roja). Todas las longitudes de onda comprendidas entre
este intervalo pertenecen al espectro visible. Cada color especifico de la luz
visible: rojo, naranja, amarillo, verde, azul, índigo y violeta, tiene una
longitud de onda y frecuencia diferentes. La mezcla de todas las longitudes de
onda de la luz visible da por resultado la luz blanca.
-Radiación ultravioleta: ‘’Luz negra’’
-Radiación ultravioleta: ‘’Luz negra’’
Tiene longitudes de onda
más corta que la luz visible, mostrada en la imagen anterior. Existen ‘’UV
cercano’’ y ‘’UV lejano’’ que son las longitudes de onda cerca y lejos
respectivamente de la luz visible. Se emplean tres categorías (UV-A / UV-B /
UV-C) indicando las longitudes de onda largas, medianas y cortas. Nuestra
atmosfera elimina por filtración las longitudes de onda UV-C.
Lo que se conoce como ‘’luz negra’’ es en realidad UV (principalmente UV cercano). Cuando la radiación UV incide en ciertas rocas o ciertos tipos de pintura, los objetos presentan fluorescencia; Parecen despedir luz propia al ser bombardeados por los rayos UV. Esto sucede cuando los electrones de los átomos de un material absorben rayos UV y luego despiden esta energía emitiendo luz visible de menor energía. La luz emitida hace que el material parezca resplandecer.
-Radiación infrarroja (IR)
Lo que habitualmente
conocemos como energía radiante es en realidad Radiación IR. Los rayos IR
tienen una longitud de onda demasiado larga para ser visibles al ojo humano,
pero poseen frecuencias idóneas para interactuar con las moléculas y producir
vibraciones moleculares.
- El control remoto de un
televisor o una videograbadora utiliza un haz de radiación IR.
-Microondas y radar
-Microondas y radar
Las longitudes de ondas de
las microondas y del radar son similares (1 cm). Las microondas tienen la
energía idónea para obligar a las moléculas a girar. Esta propiedad hace
posible que un horno de microondas caliente una salchicha rápidamente. Conforme
las microondas pasan a través de la salchicha, la energía de aquellas hace
girar las moléculas de agua del centro y también las de la superficie de la
salchicha. El calor que las moléculas producen al girar calienta rápidamente
toda la salchicha o lo que se quiere calentar. Como ya se ha señalado, las
frecuencias de luz visible constituyen tan solo una pequeña parte del espectro
electromagnético total.
1
CONCEPTOS RELATIVOS A LA LUZ
La
luz es una forma de energía radiante, y aunque su precisa naturaleza requiere
complejas teorías físicas, todos los fenómenos relativos a la óptica mineral
pueden ser correctamente explicados considerando exclusivamente su naturaleza
ondulatoria, así, en este programa se considerará que la luz se propaga como
consecuencia de una vibración de partículas.
En
la figura siguiente se muestra como a partir de un nivel de reposo se produce
una progresiva vibración de partículas que como consecuencia originan una onda
que se propaga en dirección perpendicular a la de vibración. Por tanto, el
resultado de la vibración de partículas adyacentes es una propagación de la
onda resultante.
La luz visible abarca tan
solo una pequeña fracción del espectro electromagnético total. Nuestros ojos perciben
longitudes de onda que van desde aproximadamente 400 nm (Luz violeta) hasta
alrededor de 750 nm (luz roja). Todas las longitudes de onda comprendidas entre
este intervalo pertenecen al espectro visible. Cada color especifico de la luz
visible: rojo, naranja, amarillo, verde, azul, índigo y violeta, tiene una
longitud de onda y frecuencia diferentes. La mezcla de todas las longitudes de
onda de la luz visible da por resultado la luz blanca.
-Radiación ultravioleta: ‘’Luz negra’’
-Radiación ultravioleta: ‘’Luz negra’’
Tiene longitudes de onda
más corta que la luz visible, mostrada en la imagen anterior. Existen ‘’UV
cercano’’ y ‘’UV lejano’’ que son las longitudes de onda cerca y lejos
respectivamente de la luz visible. Se emplean tres categorías (UV-A / UV-B /
UV-C) indicando las longitudes de onda largas, medianas y cortas. Nuestra
atmosfera elimina por filtración las longitudes de onda UV-C.
Lo que se conoce como ‘’luz negra’’ es en realidad UV (principalmente UV cercano). Cuando la radiación UV incide en ciertas rocas o ciertos tipos de pintura, los objetos presentan fluorescencia; Parecen despedir luz propia al ser bombardeados por los rayos UV. Esto sucede cuando los electrones de los átomos de un material absorben rayos UV y luego despiden esta energía emitiendo luz visible de menor energía. La luz emitida hace que el material parezca resplandecer.
-Radiación infrarroja (IR)
Lo que habitualmente
conocemos como energía radiante es en realidad Radiación IR. Los rayos IR
tienen una longitud de onda demasiado larga para ser visibles al ojo humano,
pero poseen frecuencias idóneas para interactuar con las moléculas y producir
vibraciones moleculares.
- El control remoto de un
televisor o una videograbadora utiliza un haz de radiación IR.
-Microondas y radar
-Microondas y radar
Las longitudes de ondas de
las microondas y del radar son similares (1 cm). Las microondas tienen la
energía idónea para obligar a las moléculas a girar. Esta propiedad hace
posible que un horno de microondas caliente una salchicha rápidamente. Conforme
las microondas pasan a través de la salchicha, la energía de aquellas hace
girar las moléculas de agua del centro y también las de la superficie de la
salchicha. El calor que las moléculas producen al girar calienta rápidamente
toda la salchicha o lo que se quiere calentar. Como ya se ha señalado, las
frecuencias de luz visible constituyen tan solo una pequeña parte del espectro
electromagnético total.
1
CONCEPTOS RELATIVOS A LA LUZ
La
luz es una forma de energía radiante, y aunque su precisa naturaleza requiere
complejas teorías físicas, todos los fenómenos relativos a la óptica mineral
pueden ser correctamente explicados considerando exclusivamente su naturaleza
ondulatoria, así, en este programa se considerará que la luz se propaga como
consecuencia de una vibración de partículas.
En
la figura siguiente se muestra como a partir de un nivel de reposo se produce
una progresiva vibración de partículas que como consecuencia originan una onda
que se propaga en dirección perpendicular a la de vibración. Por tanto, el
resultado de la vibración de partículas adyacentes es una propagación de la
onda resultante.
Para
la explicación de las propiedades ópticas de los cristales es importante tener
siempre en cuenta que las direcciones de vibración y de propagación son
perpendiculares. Esto es estrictamente cierto para todos los medios isótropos,
pero en determinadas condiciones de los anisótropos, el ángulo puede ser
diferente de los 90 grados, sin embargo, se puede considerar que ambas son
siempre perpendiculares (aceptar esto simplicará en gran medida las
explicaciones sin que se afecten la esencia de los conceptos). Por otra parte, es igualmente
importante recordar que la propagación es un simple resultado de la vibración y
por tanto será esta la que condicione a aquella.
A
continuación se repasará muy brevemente algunos conceptos relativos a la luz.
1.1
ONDA
Es
el movimiento sinusoidal causado por un grupo de particulas vibrando.

1.1
RAYO
Es
el camino rectilíneo seguido por la onda (camino recorrido por la luz)

1.1
LONGITUD DE ONDA
Es
la distancia entre dos puntos en fase (siendo puntos en fase aquellos que
encuentran vibrando de la misma menera, a igual distancia del nivel de reposo y
moviendose en la misma dirección).

1.1
COLOR
Cuando
decimos que “vemos”
un color, nos referimos realmente a que, según la luz que entre en nuestros
ojos, sentimos un color u otro. Es decir, llevamos el color al terreno de lo
perceptual, lo percibido. En el mundo externo a nuestro sistema visual no
existe el color; ese mundo es incoloro. La materia es incolora y la luz es
incolora.
El color sólo existe
como impresión sensorial del individuo que ve un objeto material.
La sensación “color”
es el producto conceptual elaborado por nuestro cerebro a partir de los datos
emitidos por el ojo que ve un objeto iluminado, un objeto sobre el que incide
la luz. En ese sentido, ver blanco (sensación de color
blanco) es ver todo el espectro visible, comprendido entre el infrarrojo y el
ultravioleta, ambos excluidos. Ver negro (sensación de color
negro) es no ver nada de ese espectro visible. Si en vez de referirnos a la luz
nos referimos al objeto que vemos, lo vemos blanco si ese objeto refleja todo
el espectro visible y lo vemos negro si ese objeto no refleja nada del espectro
visible. El aspecto color de un objeto recibe
el nombre de color de ese objeto.
2
CUALIDADES DE LA LUZ
La utilización de la luz en las
imágenes puede llegar a modificar los atributos o cualidades de las cosas que
se quiere representar o fotografiar, en función de lo que queramos expresar,
por lo que es muy importante conocer las cualidades de la luz para aprovechar
mejor las luces naturales, pero también poder conseguir cualquier tipo de
iluminación según los valores que queramos plasmar en cada momento en nuestra
fotografía, ya sea modificando la que ya hay o poniéndola tú toda.
Las cualidades de la luz son la
intensidad, la dirección, la calidad y el color.
-Intensidad: alta, baja.
-Dirección: frontal, lateral,
trasera, cenital, nadir.
-Calidad: dura, suave.
-Color: cálida, fría.
2.1 INTENSIDAD DE LA LUZ:
La intensidad es la cantidad de
luz que incide sobre un objeto, y es cuando se dice si está más o menos
iluminado o más o menos oscuro, se puede medir con un fotómetro.
2.2 DIRECCIÓN DE LA LUZ:
La dirección debe de entenderse
desde la posición de la cámara, no desde la posición del sujeto, es decir que
cada una de estas direcciones se refiere a como incide la luz en el sujeto pero
hay que entenderlo visto desde la posición de nuestra cámara.
2.3 CALIDAD DE LA LUZ:
Según la calidad de la luz,
podemos encontrar o crear luz dura, luz suave y todos sus matices intermedios.
La calidad afecta en cómo se van
a distribuir las luces y las sombras, en la transición o gradación entre ambas,
que la luz sea dura o suave depende del tamaño de la fuente de luz con respecto
al sujeto iluminado, una misma fuente de luz producirá en un objeto luz dura
cuando el sujeto sea más grande que la fuente de luz, y luz suave cuando el
sujeto sea más pequeño que esta, y al afectar el tamaño hay que tener en cuenta
también la distancia a la que está la fuente de luz del sujeto, el sol por
ejemplo es una fuente de luz enorme, pero al estar tan lejos es un simple
punto, por eso produce sombras duras, una ventana grande o la refracción en una
superficie grande hace luz suave en una cara por ejemplo, que es más pequeña
que la fuente de luz.
3 SISTEMA VISUAL HUMANO
El sistema visual humano (SVH) es el
encargado de convertir las ondas electromagnéticas que pertenecen al espectro
visible y que llegan hasta los ojos, en señales nerviosas que son interpretadas
por el cerebro.
3.1
EL OJO HUMANO
El ojo humano es una estructura
prácticamente esférica en la que entra la luz sólo por un pequeño agujero (como
ocurre en una cámara fotográfica). La cubierta externa del ojo es opaca y el
interior del ojo es translúcido.
3.2 LA ESCLERÓTIDA
Es la membrana más externa del ojo y es
opaca, excepto en su parte anterior donde es transparente y se llama córnea. Su
función principal es la de evitar que la luz entre en el ojo, excepto a través
de la córnea.
3.3
LA COROIDES Y EL IRIS
Se trata de una membrana muy pigmentada
y vascularizada que recubre prácticamente todo el ojo. Evita que entre luz a
través de ella y su gran irrigación sanguínea proporciona calor y alimento al
resto del ojo.
En su parte anterior tiene una expansión
muscular redonda llamada iris y en el centro hay una abertura redonda llamada
pupila. La función del iris es controlar (mediante un acto reflejo) el diámetro
de la pupila, determinando así la cantidad de luz que entra en el ojo.
3.4
EL CRISTALINO Y EL MÚSCULO
CILIAR
Es una estructura transparente en forma
de lente, formado por estratos concéntricos de células fibrosas que están
unidas al músculo ciliar.De la tensión de este músculo depende la distancia
focal del ojo. Cuando el ojo está relajado, el cristalino se redondea y el ojo
enfoca al infinito.1
Además, el cristalino se encuentra
ligeramente coloreado por una pigmentación amarilla que absorbe la luz
infrarroja y ultravioleta, que podría dañar la retina.
3.5
LA CORNEA Y EL CRISTALINO
Estas dos estructuras transparentes
funcionan como las lentes del telescopio de Galieo. Gracias al fenómeno de la
difracción de la luz, la córnea concentra la luz externa para que pase a través
de la pupila. El cristalino hace la función inversa, consiguiendo que la luz se
concentre en la fóvea.
3.6 EL HUMOR ACUOSO
El ojo posee una cámara anterior rellena
de un líquido transparente llamado humor acuoso, que es una dispersión de
albúmina en agua salada2 . En esta
cámara, detrás del iris, va alojado el cristalino.
3.7
EL HUMOR VÍTREO
El ojo posee una cámara posterior que
está ocupada por el humor vítreo, una especie de gel proteínico muy frágil3 . Dicha cámara
está rodeada por la membrana hialoide (que no se ve en la figura).
3.8 LA RETINA
Es una membrana sensible a la luz y
cubre la práctica totalidad de la coroides. La retina es realmente el entramado
nervioso formado por las células terminales de las fibras del nervio óptico. En
su parte posterior presenta una pequeña depresión llamada fóvea que es muy
importante en la visión de los detalles.
En la retina se distribuyen dos tipos de
receptores de luz llamados conos y bastones.
- Los conos son sensibles al color y se localizan
principalmente en la parte posterior del ojo (en la fóvea). Para que
funcionen el nivel de iluminación debe ser suficiente.
- Los bastones (mucho más numerosos), no son
sensibles al color y se encuentran distribuidos por toda la retina.

1
ELEMENTOS BÁSICOS DE LA FÍSICA
NUCLEAR

2
CONSTITUCION DEL ATOMO Y
MODELOS ATOMICOS
La descripción básica de la
constitución atómica, reconoce la existencia de partículas con carga eléctrica
negativa, llamados electrones, los cuales giran en diversas órbitas (niveles de
energía) alrededor de un núcleo central con carga eléctrica positiva. El átomo
en su conjunto y sin la presencia de perturbaciones externas es eléctricamente
neutro.
El núcleo lo componen los protones con
carga eléctrica positiva, y los neutrones que no poseen carga eléctrica.
El tamaño de los núcleos atómicos para
los diversos elementos está comprendidos entre una cienmilésima y una
diezmilésima del tamaño del átomo.
La cantidad de protones y de
electrones presentes en cada átomo es la misma. Esta cantidad recibe el nombre
de número atómico, y se designa por la letra "Z". A la cantidad total
de protones más neutrones presentes en un núcleo atómico se le llaman número
másico y se designa por la letra "A".
Si bien hoy en día todas las
características anteriores de la constitución atómica son bastante conocidas y
aceptadas, a través de la historia han surgido diversos modelos que han
intentado dar respuesta sobre la estructura del átomo.
1.1
EL MODELO DE THOMSON
Thomson sugiere un modelo atómico que
tomaba en cuenta la existencia del electrón, descubierto por él en 1897. Su
modelo era estático, pues suponía que los electrones estaban en reposo dentro
del átomo y que el conjunto era eléctricamente neutro. Con este modelo se
podían explicar una gran cantidad de fenómenos atómicos conocidos hasta la
fecha. Posteriormente, el descubrimiento de nuevas partículas y los
experimentos llevado a cabo por Rutherford demostró la inexactitud de tales
ideas.
1.2
EL MODELO DE RUTHERFORD
Basado en los resultados de su trabajo
que demostró la existencia del núcleo atómico, Rutherford sostiene que casi la
totalidad de la masa del átomo se concentra en un núcleo central muy diminuto
de carga eléctrica positiva. Los electrones giran alrededor del núcleo
describiendo órbitas circulares. Estos poseen una masa muy ínfima y tienen
carga eléctrica negativa. La carga eléctrica del núcleo y de los electrones se
neutraliza entre sí, provocando que el átomo sea eléctricamente neutro.
El modelo de Rutherford tuvo que ser
abandonado, pues el movimiento de los electrones suponía una pérdida continua
de energía, por lo tanto, el electrón terminaría describiendo órbitas en
espiral, precipitándose finalmente hacia el núcleo. Sin embargo, este modelo sirvió
de base para el modelo propuesto por su discípulo Neils Bohr, marcando el
inicio del estudio del núcleo atómico, por lo que a Rutherford se le conoce
como el padre de la era nuclear.
1.3
EL MODELO DE BOHR
El físico danés Niels Bohr (Premio
Nobel de Física 1922), postula que los electrones giran a grandes velocidades
alrededor del núcleo atómico. Los electrones se disponen en diversas órbitas
circulares, las cuales determinan diferentes niveles de energía. El electrón
puede acceder a un nivel de energía superior, para lo cual necesita
"absorber" energía. Para volver a su nivel de energía original es
necesario que el electrón emita la energía absorbida (por ejemplo en forma de
radiación). Este modelo, si bien se ha perfeccionado con el tiempo, ha servido
de base a la moderna física nuclear.
1.4
MODELO MECANO - CUÁNTICO
Se inicia con los estudios del físico
francés Luis De Broglie, quién recibió el Premio Nobel de Física en 1929. Según
De Broglie, una partícula con cierta cantidad de movimiento se comporta como
una onda. En tal sentido, el electrón tiene un comportamiento dual de onda y
corpúsculo, pues tiene masa y se mueve a velocidades elevadas. Al comportarse
el electrón como una onda, es difícil conocer en forma simultánea su posición
exacta y su velocidad, por lo tanto, sólo existe la probabilidad de encontrar
un electrón en cierto momento y en una región dada en el átomo, denominando a
tales regiones como niveles de energía. La idea principal del postulado se
conoce con el nombre de Principio de Incertidumbre de Heisenberg.
2
RADIACIÓN
Y RADIOBIOLOGÍA
2.1
RADIACIONES
El fenómeno de la radiación consiste
en la propagación de energía en forma de ondas
electromagnéticas o partículas
subatómicas a
través del vacío o de un medio material.
La radiación propagada en forma de ondas
electromagnéticas (rayos UV, rayos gamma, rayos X, etc.) se llama radiación
electromagnética,
mientras que la llamada radiación
corpuscular es la radiación
transmitida en forma de partículas subatómicas (partículas α, partículas β, neutrones, etc.) que se mueven
a gran velocidad, con apreciable transporte de energía.
Si la radiación transporta energía suficiente
como para provocar ionización en el medio que
atraviesa, se dice que es una radiación ionizante. En caso contrario
se habla deradiación no
ionizante.
El carácter ionizante o no ionizante de la radiación es independiente de su
naturaleza corpuscular u ondulatoria.
Son radiaciones ionizantes los rayos X, rayos
γ, partículas α y parte del espectro de la radiación UV entre otros. Por otro
lado, radiaciones como los rayos UV y las ondas de radio, TV o de telefonía
móvil, son algunos ejemplos de radiaciones no ionizantes.
2.2
RADIOBIOLOGIA
La radiobiología es la ciencia que estudia los
fenómenos que se producen en los seres vivos tras la absorción de energía procedente de las radiaciones
ionizantes.
Las dos grandes razones que han impulsado la investigación de los efectos
biológicos de las radiaciones ionizantes son:
1.
Protección
Radiológica: Poder utilizar esas radiaciones de forma
segura en todas las aplicaciones médicas o industriales que las requieran.
2.
Radioterapia:
Utilización de las radiaciones ionizantes principalmente en neoplasias,
preservando al máximo los órganos críticos (tejido humano sano).
3
ORIGENES DE LAS RADIACIONES
IONIZANTES
Se define una radiación
como ionizante cuando al interaccionar con la materia produce la ionización de
la misma, es decir, origina partículas con carga eléctrica (iones).
El origen de estas
radiaciones es siempre atómico, pudiéndose producir tanto en el núcleo del
átomo como en los orbitales y pudiendo ser de naturaleza corpuscular
(partículas subatómicas) o electromagnética (rayos X, rayos gamma (γ)).
Las radiaciones ionizantes
de naturaleza electromagnética son similares en naturaleza física a cualquier
otra radiación electromagnética pero con una energía fotónica muy elevada
(altas frecuencias, bajas longitudes de onda) capaz de ionizar los átomos. Las
radiaciones corpusculares están constituidas por partículas subatómicas que se
mueven a velocidades próximas a la de la luz. Existen varios tipos de
radiaciones emitidas por los átomos, siendo las más frecuentes: la
desintegración, la desintegración "β", la emisión "γ' y la
emisión de rayos X y neutrones. Las características de cada radiación varían de
un tipo a otro, siendo importante considerar su capacidad de ionización y su
capacidad de penetración, que en gran parte son consecuencia de su naturaleza.
·
Radiación
a
Son núcleos de helio
cargados positivamente; tienen una energía muy elevada y muy baja capacidad de
penetración y las detiene una hoja de papel.
·
Radiación
b
Son electrones emitidos
desde el núcleo del átomo como consecuencia de la transformación de un neutrón
en un protón y un electrón.
·
Radiación
b+
Es la emisión de un
positrón, partícula de masa igual al electrón y carga positiva, como resultado
de la transformación de un protón en un neutrón y un positrón.
·
Las
radiaciones β
Tienen un nivel de energía
menor que las α y una capacidad de penetración mayor y son absorbidas por una
lámina de metal.
·
Radiación
de neutrones
Es la emisión de partículas
sin carga, de alta energía y gran capacidad de penetración. Los neutrones se
generan en los reactores nucleares y en los aceleradores de partículas, no
existiendo fuentes naturales de radiación de neutrones.
·
Radiación
g
Son radiaciones
electromagnéticas procedentes del núcleo del átomo, tienen menor nivel de
energía que las radiaciones α y β y mayor capacidad de penetración, lo que
dificulta su absorción por los apantallamientos.
·
Rayos
X
También son de naturaleza
electromagnética pero se originan en los orbitales de los átomos como
consecuencia de la acción de los electrones rápidos sobre la corteza del átomo.
3.1
INTERACCIÓN CON EL ORGANISMO.
EFECTOS BIOLÓGICOS
Desde el descubrimiento de
los rayos X y los elementos radiactivos, el estudio de los efectos biológicos de
las radiaciones ionizantes ha recibido un impulso permanente como consecuencia
de su uso cada vez mayor en medicina, ciencia e industria, así como de las
aplicaciones pacíficas y militares de la energía atómica. Como consecuencia,
los efectos biológicos de las radiaciones ionizantes se han investigado más a
fondo que los de prácticamente, cualquier otro agente ambiental.
La
energía depositada por las radiaciones ionizantes al atravesar las células
vivas da lugar a iones y radicales libres que rompen los enlaces químicos y
provocan cambios moleculares que dañan las células afectadas (fig. 2). En
principio, cualquier parte de la célula puede ser alterada por la radiación
ionizante, pero el ADN es el blanco biológico más crítico debido a la
información genética que contiene. Una dosis absorbida lo bastante elevada para
matar una célula tipo en división (2 Grays ver la definición más adelante),
sería suficiente para originar centenares de lesiones reparables en sus
moléculas de ADN. Las lesiones producidas por la radiación ionizante de
naturaleza corpuscular (protones o partículas alfa) son, en general, menos
reparables que las generadas por una radiación ionizante fotónica (rayos X o
rayos gamma). El daño en las moléculas de ADN que queda sin reparar o es mal reparado puede
manifestarse en forma de mutaciones cuya frecuencia está en relación con la
dosis recibida.
Las lesiones del aparato
genético producidas por irradiación pueden causar también cambios en el número
y la estructura de los cromosomas, modificaciones cuya frecuencia, de acuerdo
con lo observado en supervivientes de la bomba atómica y en otras poblaciones
expuestas a radiaciones ionizantes, aumenta con la dosis.
En consecuencia, el daño
biológico puede producirse en el propio individuo (efecto somático) o en
generaciones posteriores (efecto genético), y en función de la dosis recibida
los efectos pueden ser inmediatos o diferidos en el tiempo, con largos periodos
de latencia.
También es importante
considerar la diferencia entre efectos "estocásticos" y "no
estocásticos", según que la relación dosis respuesta tenga carácter
probabilístico, o bien el efecto se manifieste a partir de un determinado nivel
de dosis (0,25 Sv), llamada dosis umbral. En ambos casos la probabilidad de
efecto o el efecto aumenta con la dosis.
1.1
IRRADIACIÓN Y CONTAMINACIÓN
RADIACTIVA. EXPOSICIÓN
Se denomina irradiación a la
transferencia de energía la de un material radiactivo a otro material, sin que
sea necesario un contacto físico entre ambos, y contaminación radiactiva a la
presencia de materiales radiactivos en cualquier superficie, materia o medio,
incluyendo las personas. Es evidente que toda contaminación da origen a una
irradiación.

Irradiación externa
Se dice que hay riesgo de
irradiación externa cuando, por la naturaleza de la radiación y el tipo de
práctica, la persona sólo está expuesta mientras la fuente de radiación está
activa y no puede existir contacto directo con un material radiactivo. Es el
caso de los generadores de rayos X, los aceleradores de partículas y la
utilización o manipulación de fuentes encapsuladas.
Contaminación radiactiva
Cuando puede haber contacto
con la sustancia radiactiva y ésta puede penetrar en el organismo por cualquier
vía (respiratoria, dérmica, digestiva o parenteral) se habla de riesgo por
contaminación radiactiva. Esta situación es mucho más grave que la simple
irradiación, ya que la persona sigue estando expuesta a la radiación hasta que
se eliminen los radionucleidos por metabolismo o decaiga la actividad
radiactiva de los mismos.
Exposición
Se llama exposición al
hecho de que una persona esté sometida a la acción y los efectos de las
radiaciones ionizantes. Puede ser:
● Externa: exposición del organismo a fuentes
exteriores a él. ● Interna: exposición del organismo a fuentes interiores a él.
● Total: suma de las
exposiciones externa e interna.
● Continua: exposición
externa prolongada, o exposición interna por incorporación permanente de
radionucleidos, cuyo nivel puede variar con el tiempo.
● Única: exposición externa
de corta duración o exposición interna por incorporación de radionucleidos en
un corto periodo de tiempo.
● Global: exposición
considerada como homogénea en el cuerpo entero.
● Parcial: exposición sobre
uno o varios órganos o tejidos, sobre una parte del organismo o sobre el cuerpo
entero, considerada como no homogénea. En caso de contaminación radiactiva del
organismo humano, según que los radionucleidos estén depositados en la piel,
los cabellos o las ropas, o bien hayan penetrado en el interior del organismo,
se considera contaminación externa o contaminación interna respectivamente. La
gravedad del daño producido está en función de la actividad y el tipo de
radiaciones emitidas por los radionucleidos. Medida de las radiaciones
ionizantes
Los aparatos de detección y
medida de las radiaciones ionizantes se basan en los fenómenos de interacción
de la radiación con la materia. Teniendo en cuenta su funcionalidad, los
instrumentos de medida se pueden clasificar como detectores de radiación o
dosímetros.
1
RADIACIONES: NATURALEZA Y
PROPIEDADES
Como ya se ha dicho, los seres
vivos están expuestos a niveles bajos de radiación ionizante procedente del
sol, las rocas, el suelo, fuentes naturales del propio organismo, residuos
radiactivos de pruebas nucleares en el pasado, de ciertos productos de consumo
y de materiales radiactivos liberados desde hospitales y desde plantas
asociadas a la energía nuclear y a las de carbón.
Los trabajadores expuestos a
mayor cantidad de radiaciones son los astronautas (debido a la radiación
cósmica), el personal médico o de rayos X, los investigadores, los que trabajan
en una instalación radiactiva o nuclear. Además se recibe una exposición
adicional con cada examen de rayos X y de medicina nuclear, y la cantidad
depende del tipo y del número de exploraciones.
No se ha demostrado que la
exposición a bajos niveles de radiación ionizante del ambiente afecte la salud
de seres humanos. De hecho existen estudios que afirman que podrían ser
beneficiosas (la hipótesis de la hormesis).1 2 Sin embargo, los organismos
dedicados a la protección radiológica oficialmente utilizan la hipótesis
conservadora de que incluso en dosis muy bajas o moderadas, las radiaciones
ionizantes aumentan la probabilidad de contraer cáncer, y que esta probabilidad
aumenta con la dosis recibida (Modelo lineal sin umbral).3 4 A los efectos
producidos a estas dosis bajas se les suele llamar efectos probabilistas, estadísticos
o estocásticos.
La exposición a altas dosis de
radiación ionizante puede causar quemaduras de la piel, caída del cabello,
náuseas, enfermedades y la muerte. Los efectos dependerán de la cantidad de
radiación ionizante recibida y de la duración de la irradiación, y de factores
personales tales como el sexo, edad a la que se expuso, y del estado de salud y
nutrición. Aumentar la dosis produce efectos más graves.
1
RADIOACTIVIDAD
La radioactividad puede ser natural o artificial.
· En la radioactividad natural, la sustancia ya la posee en
el estado natural.
· En la radioactividad artificial,
la radioactividad le ha sido inducida por irradiación.
Un radionucleido es el conjunto de los núcleos
radioactivos de una misma especie. Todos los núcleos radioactivos que
forman un radionucleido tienen una radiactividad bien definida, común a todos
ellos, que los identifica; de la misma forma que un tipo de reacción
química identifica los elementos que participan.
1.1
Nucleidos radioactivos naturales
En la naturaleza
se encuentran unos 300 nucleidos diferentes, de los cuales 25 son radioactivos
con un período suficientemente largo para que haya aún hoy en día; otros
35 tienen un período mucho más corto y se crean y se desintegran continuamente
en las series radiactivas.
1.2
Nucleidos radioactivos artificiales
Han
sido creados e identificados más de 1000 radionucleidos artificiales. Las
series radioactivas reciben el nombre del nucleido padre de períodos más
largo. Hay cuatro. Tres de estas series radioactivas son naturales:
la del torio, la del uranio y la del actinio, que terminan en sus propios
isótopos estables del plomo. Estos isótopos tienen
respectivamente los números de masa 208, 206 y 207. Respecto a la serie del
neptunio, como los radionucleidos que la componen tienen un período corto
comparado con la duración de las eras geológicas, no se encuentra en la
naturaleza y ha sido obtenida artificialmente. El último nucleido de esta
serie es el isótopo 209
del bismuto.
1.3
Origen de la radioactividad
La
radioactividad fue descubierta en 1896 por Antonie-Henri Becquerel,
el cual, al hacer estudios sobre la fosforescencia de las sustancias, observó
que un mineral de uranio era capaz de velar unas placas fotográficas que
eran guardadas a su lado.
1
RAYOS X

Probablemente se sometió a un
examen con rayos X en alguna parte del cuerpo. Los profesionales de la salud
los usan para observar los huesos rotos, problemas en los pulmones y el
abdomen, caries dentales y muchos otros problemas. Por ejemplo, las mamografías
usan rayos X para observar tumores o áreas sospechosas en los senos.
La tecnología de rayos X usa
radiación electromagnética para producir imágenes. La imagen se registra en una
película o placa llamada radiografía. Las partes del cuerpo aparecen claras u
oscuras debido a las distintas tasas de velocidad a las que los tejidos
absorben los rayos X. El calcio de los huesos lo hace al máximo, por lo que los
huesos se ven blancos en la radiografía.
La grasa y otros tejidos blandos
absorben menos y se ven grises. El aire absorbe menos, por lo que los pulmones
se ven negros.
Los estudios con rayos X no
duelen, son rápidos y fáciles. La cantidad de exposición a la radiación que
recibe durante un estudio con rayos X es pequeña.
No hay comentarios.:
Publicar un comentario